
铝胁迫对海莲幼苗中某些金属元素累积的影响
2010-03-15 10:12杨盛昌
海洋科学 2010年8期
马 丽,杨盛昌
(1.厦门大学 生命科学学院 滨海湿地生态系统教育部重点实验室,福建厦门 361005; 2.河北承德石油高等专科学校,河北 承德 067000)
Al占地壳组成的8%,在自然界中一般以氧化物和硅酸盐(如石英、云母等)的形式存在,是地壳中含量最丰富的金属元素。与其在地壳中的丰富含量对比,海洋中Al的含量在1pg/L 以下,这可能是与硅藻属植物对Al的富集作用有关; 海水中Al多以Al3+形式存在,自由的Al3+以羟化物的形式沉淀,除了在活火山口和矾矿地带,自然水源中Al的富集并不显著[1]。
Al以自然的固态形式存在时,不会对植被和环境产生毒害和污染。酸雨是金属离子释放的主要途径,随着世界范围内酸雨的影响,铝、汞、铅等金属离子从矿床流失进入淡水,以无机离子态存在的 Al在土壤中大量渗出,导致土壤富 Al化,成为酸性土壤植物生产力的主要限制因素,直接导致了农作物减产和森林面积退化[1~3]。通过长期的血液透析等研究手段发现,Al对动物和人的健康也产生严重毒害作用,如抵抗维生素 D的骨软化、小红细胞贫血、帕金森症等疾病[1]。因此,Al盐毒害以及植物的耐Al机制成为备受关注的研究热点之一。
红树林是分布在热带亚热带海岸潮间带的木本植物群落。红树植物具有耐盐和抗水淹等生理生态学特性[4,5],对汞、镉、铅等重金属元素有较强的富集作用和耐受特性[6,7],在治理环境污染上有一定应用前景。国内外学者对红树林开展了大量的研究,但有关Al胁迫对红树植物影响的报道较少。作者以中国常见的红树林树种之一海莲(Brugiera sexangula)为材料,研究了Al胁迫对海莲幼苗中Na、Mg、K、Ca、Mn、Fe、Cu、Zn等金属离子的吸收累积和离子间相互作用的影响,并对可能的缓解Al毒害的离子机制进行了探讨。
1 材料与方法
1.1 材料培养及Al处理
海莲成熟胚轴采于海南省文昌市清澜港红树林自然保护区,平均长度8.23 cm±1.04 cm、平均质量12.2 g±0.68 g。将海莲胚轴于厦门大学生命科学学院温室内进行砂培。沙砾粒径2~4 mm,经自来水反复冲洗后装入塑料网盆中,每盆质量约2.5 kg,随机分组,每盆种植海莲胚轴 5~6棵。培养液采用Hoagland’s液体培养基,并混合3‰NaCl以保证海莲生长的生理需盐,每隔 5d更换一次培养基,每天补充因蒸发损失的水分。
1个月后采用氯化铝溶液进行铝胁迫处理,铝(Al3+)浓度分别为10、25、50 mmol/L,以未添加铝盐的培养基作为对照。每一处理重复4次,随培养基更换分批次添加氯化铝,培养期为2个月。
1.2 试验方法
1.2.1 仪器和试剂
主要仪器为微波消解系统(MARS5)、电感耦合等离子体原子发射光谱分析系统(ICP-MS 美国 PE公司)。
元素标准样、硝酸、双氧水等药品试剂均为优级纯,超纯水配置相关溶液。
1.2.2 样品的预处理
收获海莲幼苗,并按照叶、茎(含胚轴)、根分组,85℃过夜烘干,然后研磨成干粉样。准确称取海莲各组分干粉样品 0.100 g,装入已经用硝酸清洗过的消化罐内杯底部,加入4 mL HNO3,使酸完全浸没样品,再加入1 mL H2O2,盖上密封盖,拧紧外壳压盖后放入微波炉中进行消解[8]。消解结束时降温到 30℃左右,从微波炉中取出消化罐,将溶液转移至 100 mL容量瓶中,用超纯水定容,摇匀备用(置于 4℃冰箱中)。
1.2.3 标准曲线及样品测定
配置所测金属元素标准溶液并用超纯水稀释成不同浓度,以超纯水为空白,测定并绘制标准曲线。测定不同浓度Al处理后的海莲各组分样品Na、Mg、Al、K、Ca、Mn、Fe、Cu、Zn 的含量,每个样品重复3次以上。
1.2.4 数据统计分析
采用单因素方差分析法(One-way ANOVA)对实验数据进行统计分析。所用软件为SPSS11.0。
2 结果与分析
2.1 Al在海莲幼苗各器官中的积累
如图1所示,铝盐处理下,海莲叶片、茎、根部对Al的积累量均呈现增加趋势,以根部对Al的积累量增加最为显著,增幅为 329.63%~521.43%。因此,在Al胁迫下,海莲根部是积累Al的最重要部位。根对 Al的大量富集,使植株在一定浓度范围内的 Al胁迫下,减少了 Al向地上部的运输,从而缓解对叶片的伤害。
海莲幼苗各器官对Al的富集系数见表1。从各个器官来看,同一器官对Al的富集系数均随着培养液中铝盐浓度的提高而呈下降趋势。在同一铝盐浓度下,根部的富集系数最大,茎部最小,其中,在10 mmol/LAl处理下,根对Al的富集系数是同质量浓度下茎的4.2倍,叶的2.9倍。
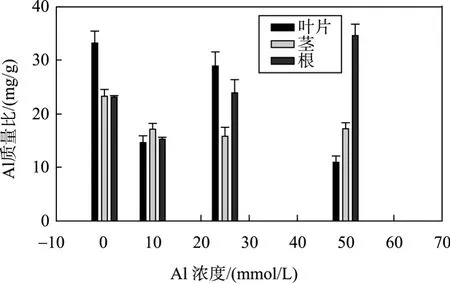
图1 Al在海莲幼苗各器官的积累Fig.1 Accumulations of Al in Brugiera sexangula seedlings

表1 海莲幼苗对Al的富集系数Tab.1 Al3+ enrichment coefficients of Brugiera sexangula seedlings
2.2 Al对海莲中Na、K、Mg、Ca积累的影响
如图2所示,Al处理下,Na在海莲根部的积累量较对照有显著增加,而在叶片和茎中的含量变化不显著。
K在海莲各器官中的积累量变化不一致。与对照组相比,在10 mmol/L Al处理下根对K的积累有显著增加,而更高浓度处理时,K在根中的积累量变化不显著。当Al浓度增至50 mmol/L时,茎中K的积累量显著增加,而25 mmol/L和50 mmol/L处理时,叶片中K的含量较对照下降。
Mg在海莲各器官中的积累量变化也不一致,在10 mmol/L和25 mmol/L Al处理下,Mg在根中的积累较对照有所下降,在这两个浓度处理下,茎和叶对Mg的积累较对照无显著变化; 在50 mmol/L Al处理下,Mg在茎中的积累反而较对照有显著增加。
Ca在海莲各器官的积累情况为:在 25 mmol/L和50 mmol/L处理时,海莲叶片对Ca的积累显著下降,但在根部的积累明显增加,茎中变化不大。
表2结果显示,10 mmol/L Al处理下,根部的Na/K降低,可能是Al刺激了海莲根系对K的选择性吸收所致。但在25 mmol/L和50 mmol/L两个浓度下,根部的Na/K显著增加,这可能与海莲对Na的吸收增强有关。
随着Al盐浓度增加,Al/Ca较对照组显著提高。因此,在高Al浓度下,Ca对Al的缓解作用明显下降。

表2 海莲幼苗各器官中Na/K、Al/CaTab.2 Na/K and Al/Ca in Brugiera sexangula seedlings

图2 Al对海莲幼苗各器官中Na、Mg、K、Ca的积累的影响Fig.2 Accumulations of Na,Mg,K,Ca in Brugiera sexangula seedlings
2.3 Al对海莲幼苗各器官中Mn、Fe、Cu、Zn四种微量元素积累的影响
Al对海莲幼苗各器官中Mn、Fe、Cu、Zn四种微量元素积累的影响见图3。Al处理下,Mn在海莲植株各部位的积累量较对照减少。结果说明,Al抑制了根对Mn的吸收积累,相应减少了海莲地上部位对Mn的积累。
Fe在海莲叶片、茎中的积累随Al浓度的增加出现减少趋势。表明Al抑制了 Fe在海莲幼苗地上部位的积累。根系仅在50 mmol/L Al处理下对Fe的积累受到显著抑制,较对照减少了70.1%。
海莲根部对 Cu的积累量较对照显著减少; 50 mmol/L Al处理下,茎对Cu的积累则显著增加; 在叶中 Cu的积累则没有一定的规律性,这可能与海莲对重金属的富集部位的特异性有关。
Al处理下,Zn在海莲茎中的积累较对照有显著增加,叶片和根中对Zn的积累则缺乏规律性。
3 讨论
Al胁迫下海莲幼苗各器官中金属元素的积累结果表明,Al在海莲幼苗各器官的含量均随着Al处理液浓度的提高而增加; 尤其在根中的增加最为显著。从 Al在植株的分配来看,两个月的胁迫处理期中,大量的Al在海莲根部沉积; 随Al浓度的增加,根对Al的分配比例增加,可能因为海莲根部对Al的固定作用,吸收后还没有运输到相应部位,相应地降低了茎和叶中Al的相对含量。根对Al的富集,减轻了Al对茎、叶生长的毒害,也是植物对Al胁迫的一种适应机制。

图3 Al对海莲幼苗各器官中Mn、Fe、Cu、Zn的积累的影响Fig.3 Accumulations of Mn,Fe,Cu,Zn in Brugiera sexangula seedlings
Al胁迫下,海莲根部对Na、K离子吸收有所增加; 虽然根对Mg的吸收有所下降,但Mg向地上部位的运输并未受到抑制,这与李海生等[9]在研究 Al对茶苗的影响中的报道一致,这说明Al对钠、钾、镁的吸收和运输的负作用不大。
海莲植株 Ca/Al比下降,钙的积累在叶片相对减少,这可能是 Al3+阻塞了 Ca2+离子通道所致; 海莲根中Ca的含量在25 mmol/L和50 mmol/L浓度下,较对照有所增加,可能是根加大了对 Ca的主动吸收以暂时缓解Al毒害。
Mg是植物光合作用和呼吸作用中各种磷酸变位酶和磷酸激酶的活化因子,在 DNA和 RNA的合成过程中也发挥重要的活化作用。Al的原初毒害是根原生质膜 Ca2+通道受阻,导致根尖细胞 Ca2+净吸收下降,原生质 Ca2+缺乏,Ca2+平衡破坏,进而影响细胞结构和功能,导致 Mg2+在根系阳离子交换量的饱和度下降,Mg2+的吸收下降,可能是 Al3+竞争了Mg2+在根质外体上的结合位点[10]。
Ca是植物必须营养元素,对维持细胞壁、细胞膜的稳定性,调控植物体内酶和阴阳离子平衡具有重要作用。而且作为植物细胞中的第二信使,Ca2+感受、传递和响应环境信号的变化,可能直接调控部分抗氧化酶活力[11]。Al毒害干扰了Ca2+的吸收与平衡,导致了膜脂过氧化,降低了 H+泵活性,抑制了 Ca2+-ATP的活性。已有的研究表明,钙能减轻酸雨[12]、盐胁迫[13]和Al毒[14]等多种胁迫对植物的毒害作用。
Al处理下,Mn、Fe、Cu、Zn在海莲某些器官中的积累有不同程度的减少,但是由于植物对这些微量元素的需求量小,因此受 Al毒的影响很小,表明海莲根系对Al毒胁迫有较强的适应性。
Fe是植物生长必需的营养元素,在维持细胞内物质代谢、蛋白质合成、叶绿体发育、酶活性等生理功能方面发挥重要作用。照光叶绿体中,捕光色素的光氧化反应能产生大量氧自由基,如果缺乏清除机制,会破坏蛋白质和核酸; SOD作为一种保护酶,起重要的清除氧自由基的作用。铁也是细胞色素蛋白,铁氧还蛋白,铁硫基蛋白中不可缺少的元素,缺铁将导致光合作用系统Ⅱ的反应中心蛋白质含量显著下降[15]。
Mn作为 SOD酶的结合因子也会干扰酶活性的变化; 在糖酵解和三羧酸循环中作为酶活化剂可以提高呼吸速率,还是硝酸还原酶的活化剂。
研究结果表明,Al抑制了海莲对铁和锰的吸收和运输,可能导致SOD、CAT活性及某些功能蛋白等一系列生理生化指标受到不同程度的影响。
Cu和 Zn是植物生长所需的微量营养元素,是多种氧化酶活性的核心,参与电子的接受与传递,在植物体内的氧化还原反应中起重要作用,与叶绿素的形成以及碳水化合物、蛋白质的合成有密切关系,还能提高植物的呼吸强度[16]。
过量的Cu和Zn也是环境污染元素,在不同浓度Al处理下,随着基质中金属元素含量的变化,海莲根系对Cu、Zn的富集力也呈不同的变化趋势,说明同一植物在同一基质中对不同金属元素的滞留能力不同。余国泰等[17]发现Al毒害对小麦吸收Zn的影响不大,陈文荣等[10]研究Al对荞麦吸收运输营养元素时,也发现Al对Cu、Zn等微量元素的影响很小 ;本研究中,Al抑制海莲根部对Cu的吸收,而根对Zn的吸收受Al的影响变化不规律的相关机制有待进行更深入的探讨。
[1]Martin R B.Aluminum:A neurotoxin product of acid rain [J].Acc Chem Res,1994,27:204-210.
[2]Delhaize E,Ryan P R.Uptake on environmental stress:aluminum toxicity and tolerance in plants [J].Plant Physiol,1995,107:2315-2321.
[3]孔繁翔,桑伟莲,蒋新,等.铝对植物毒害及植物抗铝作用机理[J].生态学报,2000,20(5):855-862.
[4]张宜辉,王文卿,林鹏.短时间和长时间盐度对木榄幼苗生长及叶片膜脂过氧化作用的研究[J].水生生物学报,2004,28(2):186-190.
[5]陈鹭真,王文卿,林鹏.潮汐淹水时间对秋茄幼苗生长的影响[J].海洋学报,2005,27(2):141-147.
[6]林鹏,陈荣华.九龙江口红树林对汞的循环和净化作用[J].海洋学报,1989,11(2):242-247.
[7]杨盛昌,吴琦.Cd对桐花树幼苗生长及某些生理特性的影响[J].海洋环境科学,2003,22(1):38-42.
[8]张昕,赵永和.微波消解 ICP-AES法测定植株金属元素含量的研究[J].河南科学,2005,23(5):675-678.
[9]李海生,张志权.不同铝水平下茶对铝及矿质养分的吸收与累积[J].生态环境,2007,16(1):186-190.
[10]陈文荣,刘鹏,黄朝表,等.铝对荞麦铝和其他营养元素运输的影响[J].水土保持学报,2006,20(3):173-176.
[11]Gong M,Chen S N,Song Y Q,et al.Effect of calcium and calmodulin on intrinsic heat tolerance in relation to antioxidant system in maize seedlings [J].Plant Physil,1997,24:371 - 379.
[12]何龙飞,沈振国,刘友良.铝胁迫下钙对小麦根系细胞质膜ATP 酶活性和膜脂组成的效应[J].中国农业科学,2003,36(10):1139-1142.
[13]罗虹,刘鹏,李淑.硅、钙对水土保持植物荞麦铝毒的缓解效应[J].水土保持学报,2005,19(3):101-104.
[14]沈宏,严小龙.铝对植物的毒害和植物抗铝毒机理及其影响因素[J].土壤通报,2001,32(6):281-285.
[15]Green R M,Geider R J,Falkowski P G.Effect of iron-limitation on photosynthesis in a marine diatom [J].Limnol Oceanogr,1991,36:1772-1782.
[16]李红敬,谢素霞.铜对紫背浮萍的影响[J].广西植物,2006,23(4):362-366.
[17]余国泰,秦遂初.有机肥缓解小麦铝毒效果的研究[J].植物营养与肥料学报,1998,4(1):57- 62.
猜你喜欢
科学导报·学术(2020年19期)2020-07-09
金属加工(热加工)(2020年12期)2020-02-06
中成药(2018年11期)2018-11-24
上海农业学报(2017年3期)2017-04-10
汉语世界(2016年3期)2016-11-16
腹腔镜外科杂志(2016年11期)2016-06-01
汉语世界(The World of Chinese)(2016年3期)2016-01-10
磁共振成像(2015年9期)2015-12-26
实用手外科杂志(2015年2期)2015-08-28
养殖与饲料(2014年10期)2014-02-28
