
野生大豆与栽培大豆的同工酶分析*
2010-06-04 05:12韩琳娜周凤琴
大豆科技 2010年1期
韩琳娜,周凤琴
(山东中医药大学药学院,济南250355)
大豆(Glycine max(Linn.)Merr.)是我国主要的油料作物之一,已有五千年栽培历史,通常被认为是由野生大豆(Glycine soja Sieb.et Zucc.)驯化而来[1]。野生大豆具有耐盐碱、抗寒、抗病等许多优良性状[2,3],是重要的野生资源植物,蕴藏着丰富的遗传多样性。野生大豆在黄河三角洲有大面积分布,垦利县大汶流最多,常形成单一的野生大豆群落[4]。近年来,对野生大豆和栽培大豆的比较研究多见于有效成分比较[5]、种子贮藏蛋白研究[6]、生化及分子生物学分析[7、8]、抗逆性研究[9、10],而黄河三角洲野生大豆与栽培大豆同工酶的比较分析尚未见报道。
同工酶是基因表达的产物,特异的基因表达产生特异的同工酶酶谱。目前同工酶已被广泛的应用于植物系统演化研究、种质资源的鉴定及亲缘关系的研究等方面,都取得了良好的效果[11]。本实验利用聚丙烯酰胺凝胶电泳技术分析东营市河口区的野生大豆和山东4个栽培大豆的过氧化物酶和酯酶同工酶酶谱特征,从而有助于进一步了解野生大豆与栽培大豆之间的亲缘关系及进化,为野生大豆资源的开发利用提供一定的理论依据。
1 材料和方法
1.1 材料
野生大豆采自黄河三角洲东营市河口区滨海湿地,栽培大豆鲁豆1、鲁豆10、河豆12和中黄20均购自山东省农业科学院。上述材料的种子播种于药用植物园。所有材料经山东中医药大学周凤琴教授鉴定。
1.2 方法
1.2.1 样品处理 种子萌发后,同一时间取其新鲜幼嫩叶片0.5g,加1mL提样缓冲液和少量石英砂,在冰浴中研成匀浆,以2mL提取液分几次洗入离心管,然后在4℃以8000r/min离心10min,取上清液,加入等量40%蔗糖溶液,摇匀,置于-20℃的冰箱中保存备用。
1.2.2 电泳 采用双面垂直板聚丙烯酰胺凝胶电泳,2种同工酶分离胶浓度均为7.5%,浓缩胶浓度均为3%。点样30uL,0.1%溴酚蓝作为前沿指示剂,pH 8.3Tris-甘氨酸缓冲液作为电极缓冲液。于4℃冰箱中220V电压条件下电泳,待指示剂距玻璃板末端1.0cm处停止电泳。
1.2.3 染色 POD同工酶采用醋酸联苯胺法(4mL联苯胺贮液+18mL H2O+2滴 H2O2)染色,常温下染色5min左右,水冲洗后照相。EST同工酶采用醋酸奈酯-坚牢蓝RR盐法(α-醋酸奈酯25mg,β-醋酸奈酯25mg,坚牢蓝RR 50mg+少许丙酮+75mL pH 6.5的磷酸缓冲液)染色,常温下染色25min左右,水冲洗后照相。
1.2.4 数据分析 依据电泳照片,记录数据,求出相对迁移率和相似系数。酶带的相对迁移率(Rf)=酶带的迁移距离/溴酚蓝的迁移距离;同工酶酶谱间相似系数c=2w/(a+b),其中c为酶谱间相似系数,w为A和B两个分类群相同的酶带数,a为分类群A在酶谱中的酶带数,b为分类群B在酶谱中的酶带数。
2 结果与分析
2.1 POD同工酶
野生大豆与栽培大豆的过氧化物酶同工酶谱见图1。
从图1可知,野生大豆与栽培大豆的过氧化物同工酶谱型存在差异。栽培大豆共有9条酶带,而野生大豆共有6条酶带。根据泳动速率,酶谱可明显分为2个部分,即慢区(S)和快区(F)。慢区Rf在0.021~0.277,快区 Rf在 0.447~0.534(见表 1)。其中S1、S2为强带,其余多为弱带。S1、S2、S3、S5、S6、S7和F1、F2、F3为栽培大豆所有,S1、S2、S4、S6、S7和 F1为野大豆所有,即 S1、S2、S6、S7和 F1是大豆的特征谱带,S4是野生大豆独有的,而 S3、S5、F2、F3是栽培大豆独有的谱带。

表1 过氧化物酶带的迁移率

图1 野生大豆与栽培大豆过氧化物酶同工酶图谱
2.2 EST同工酶
野生大豆与栽培大豆的酯酶同工酶谱见图2。

表2 酯酶带的迁移率
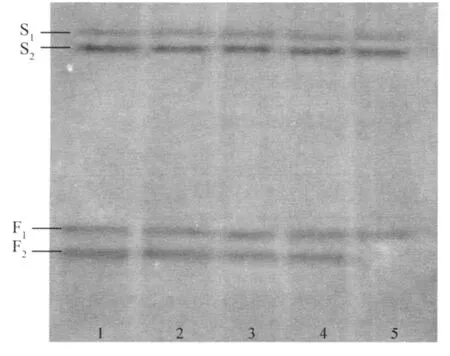
图2 野生大豆与栽培大豆酯酶同工酶图谱
从图2可以看出,野生大豆与栽培大豆的酯酶同工酶谱也存在不同。栽培大豆在慢区和快区各有2条 带 , 分 别 为 S1、S2和 F1、F2。Rf值 分 别 为0.146,0.188,0.583和 0.646(见表 2),而野生大豆共有3条酶带。S1、S2、F1为野生大豆和栽培大豆所共有的弱带,F2是栽培大豆独有的谱带,而野生大豆没有。
2.3 基于同工酶酶谱的相似系数分析
酶谱相似系数的大小表示品种亲缘关系的远近。本实验中,经分析可得,野生大豆与栽培大豆过氧化物酶同工酶酶谱的相似系数为0.667,酯酶同工酶酶谱的相似系数为0.714。从这两种酶谱分析来看,野生大豆与栽培大豆之间的亲缘关系比较近。
3 结论
野生大豆和栽培大豆不仅在植物外部形态和生境上存在差别,而且同工酶酶谱也存在明显差别。在本次实验中,野生大豆与栽培大豆过氧化物同工酶谱存在比较明显的差异,酶带分离效果较好。酯酶同工酶也存在区别,但效果不如过氧化物酶同工酶。过氧化物同工酶是由单基因决定的同工酶,它在一定程度上能较好的反映不同种源间的遗传差异和亲缘关系,而且利用其进行物种的辅助鉴定,是一种较为方便、有效的手段。因此过氧化物酶同工酶谱可以作为野生大豆和栽培大豆进化和类型鉴定的参考依据。但遗传差异不仅仅体现在一两种酶上,如果能同时对更多的同工酶进行分析,则更能全面的反映遗传差异。
野生大豆与栽培大豆的过氧化物酶和酯酶同工酶谱虽然都存在差异,但更多的是相似性,二者之间的酶谱相似系数说明二者亲缘关系较近,但也有各自的遗传基础。
[1]林镕,俞德浚,吴征镒,等.中国植物志[M].北京:科学出版社,2005.
[2]Doyle J J,Doyle J L.Ribosomalgene variation in soybean(Glycine)and its relatives [J].Theoretical and Applied Genetics,1985,70(4):369-374.
[3]於丙军,罗庆云,曹爱忠,等.栽培大豆和野生大豆耐盐性及离子效应的比较[J]植物资源与环境学报,2001,101:25-29.
[4]吴立新.野大豆栽培技术与利用[J].四川草原,2004,106(9):59-60.
[5]周三,关崎春雄,岳旺,等.野生大豆、黑豆和大豆的异黄酮类成分比较[J].2008,27(2):315-319.
[6]李春梅,杨守萍,盖钧镒,等.野生大豆与栽培大豆种子差异蛋白质组学研究[J].生物化学与生物物理进展,2007,34,(12):1296-1302.
[7]刘润堂,贾炜珑,温琪汾,等.栽培大豆、野生大豆和半野生大豆酯酶同工酶的研究[J].山西农业大学学报,1995,15(3):235-237.
[8]赵洪锟,王玉民,李启云,等.中国不同纬度野生大豆和栽培大豆 SSR 分析[J].大豆科学,2001,20(3):172-176.
[9]付畅,关旸,徐娜.盐胁迫对野生和栽培大豆中抗氧化酶活性的影响[J].大豆科学,2007,26(2):144-148.
[10]於丙军,罗庆云,曹爱忠.栽培大豆和野生大豆耐盐性及离子效应的比较[J].植物资源与环境学报,2001,10(2):25-29.
[11]刘海学,王罡,季静,刘鹏,等.16个向日葵品种过氧化物酶同工酶分析[J].中国油料作物学报,2007,29(2):64-67.
猜你喜欢
作物学报(2022年2期)2022-11-06
智慧农业导刊(2022年17期)2022-09-16
河北科技师范学院学报(2022年2期)2022-08-26
中国现代中药(2021年9期)2021-11-16
宁夏医学杂志(2020年3期)2021-01-21
信阳农林学院学报(2019年2期)2019-01-12
天然产物研究与开发(2018年10期)2018-11-06
广东农业科学(2017年5期)2017-08-29
辽宁农业科学(2017年1期)2017-03-16
现代检验医学杂志(2016年5期)2016-08-20
