
番茄茎基腐病病原菌的生物学特性
2010-06-12 03:10刘志恒马家瑞韩翔宇
植物保护 2010年2期
刘志恒, 马家瑞, 杨 红, 韩翔宇, 吕 彬, 王 悦
(沈阳农业大学植物保护学院,沈阳 110161)
近年来,随着农业产业结构的调整,早春大拱棚或温室种植番茄的面积逐年扩大[1],加之全球气候变暖,早春番茄生产上茎基腐病的发生有加重趋势[2],若该病在番茄生产上严重发生,将会对番茄生产构成严重威胁。该病害在国内外研究资料中报道很少,特别是系统性研究报道更为少见。为明确番茄茎基腐病病原菌的生物学特性,探讨病菌越冬侵染特点,本试验对采集分离的番茄茎基腐病病菌的生物学特性进行了研究。
1 材料与方法
1.1 供试材料
番茄茎基腐病害标样采自辽宁省沈阳市和抚顺市。
1.2 病害症状描述
根据番茄茎基腐病茎基部危害的症状特点进行描述记载[3]。
1.3 病原菌鉴定
按常规方法对番茄茎基腐病标样进行分离、培养、鉴定[4],描述记载病原菌形态。
1.4 病菌生物学特性测定
1.4.1 菌丝生长和菌核产生特性测定
1.4.1.1 不同培养基的影响
选用 PDA 、PSA 、2%水琼脂[5]、Richard、番茄煎汁[6]、燕麦片和番茄燕麦片7种培养基,移植直径8mm的菌饼,25℃恒温培养。设3次重复。间隔24h采用十字交叉法测定菌落直径,3d后绘制菌丝生长速度曲线,进行方差分析和多重比较[7]。20d后记录菌核数量。
番茄煎汁琼脂培养基:番茄汁150mL,琼脂粉12g,水 1000mL。
燕麦片琼脂培养基:燕麦片30g,琼脂粉12g,水1000mL。
番茄燕麦片琼脂培养基:番茄汁150mL,燕麦片30g,琼脂粉12g,水1000mL。
1.4.1.2 不同碳源的影响
PDA为基础培养基,以果糖、木糖、甘露糖、半乳糖、蔗糖、麦芽糖、乳糖、山梨糖、淀粉及糊精等量置换葡萄糖,制成 11种培养基。菌饼应用同上,25℃恒温培养[8]。重复 3次。菌落直径测定、记载、统计方法以及菌核产生记录等均同上。
1.4.1.3 不同氮源的影响
PA为基础培养基,按0.5%分别加入硝酸钾、硫酸铵、磷酸二氢铵、氯化铵、硝酸钠、蛋白胨、脲、精氨酸、胱氨酸及丙氨酸,制成11种培养基。菌饼应用同上,25℃恒温培养[8]。重复3次。菌落直径测定、记载、统计方法以及菌核产生记录等均同上。
1.4.1.4 不同温度的影响
将直径8mm菌饼移植于PDA平板培养基上,温度设置 0、2、5、10、15、20、25、30、32 、35 ℃共 10 个梯度。重复3次。间隔24h十字交叉测量菌落直径,3d后绘制菌丝生长速度曲线,进行方差分析和多重比较[7]。并于20d后记录菌核数量。
1.4.1.5 不同pH的影响
采用 PDA 平板培养基,pH 设置 2、3、4、5、6、7、8、9、10、11共10个梯度(用0.1%的 NaOH 和 HCl溶液调配)。菌饼应用同上,25℃下恒温培养。重复3次。菌落直径测定、记载、统计方法以及菌核产生记录等均同上。
1.4.1.6 不同光照的影响
将直径9mm的菌饼接种于PDA平板上,设置光照24h、黑暗 24h、L∥D=12h∥12h共 3种处理,25℃恒温培养。重复3次。菌落直径测定、记载、统计方法以及菌核产生记录等均同上。
1.4.2 菌丝致死温度测定
将直径8mm的菌饼置于装有无菌水的试管中,试管分别置于 47、48、49 、50、51、52、53 、54、55 ℃的恒温水浴锅中处理10min,迅速冷却后移植于PDA平板培养基中央,每皿3块,25℃恒温培养。每处理3次重复。3d后观察菌丝生长状况,确定菌丝致死温度。
1.4.3 菌核萌发条件测定
1.4.3.1 不同碳源的影响
采用菌丝生长和菌核产生特性测定所用的11种培养基。分别植入菌核50粒/皿,25℃恒温培养。重复3次。3d后观察菌核萌发情况,每处理随机检查100粒菌核,计算比较菌核萌发率。
1.4.3.2 不同温度的影响
采用 PDA 平板培养基 ,设置 0、5、10、15、20、25、30℃共7个温度梯度,分别移入菌核 50粒/皿。重复3次。菌核萌发观察及计算比较方法同上。
1.4.3.3 不同pH的影响
同前述方法调节 pH,设置 2、3、4、5、6 、7 、8、9、10共9个梯度。分别移入菌核 50粒/皿。重复 3次。菌核萌发观察及计算比较方法同上。
1.4.4 菌核致死温度测定
将菌核置于装有无菌水的试管中,分别置于57、58、59、60、61、62 ℃的 恒温水浴锅中处 理10min,迅速冷却后将菌核移植于PDA平板培养基,每皿4粒,25℃恒温培养。设3次重复。3d后根据菌核是否萌发生成菌丝,确定菌核致死温度。
2 结果与分析
2.1 病害症状描述
番茄茎基腐病主要危害大苗、定植后番茄的茎基部,有时延及主侧根。发病部位初呈暗褐色,继而绕茎基部或根茎处扩展,引起皮层腐烂;导致地上部叶片变黄,严重时果实膨大后植株因养分供应不足渐萎蔫枯死,发病后期在病部表面常形成黑褐色、大小不一的菌核。
2.2 病原菌形态描述
番茄茎基腐病的病原菌为立枯丝核菌(Rhizoctoniasolani),属真菌界,无性型真菌,丝孢纲,无孢目,丝核菌属。
人工培养条件下,约15d左右,菌丝彼此交织纠集形成褐色或黑褐色的菌核,球形、长粒状或不规则形,大小2.3mm×1.5mm;菌核表面粗糙,内外层颜色较一致,结构疏松。菌丝初期无色,老熟后渐为褐色,平均直径为9.6μm;菌丝分支多呈直角,近分支处形成隔膜,分支基部呈缢缩状(图1)。

图1 番茄茎基腐病病原菌形态图
2.3 病菌生物学特性测定
2.3.1 菌丝生长和菌核产生特性测定结果
2.3.1.1 不同培养基的影响
在燕麦片培养基上,菌丝生长速度显著快于其他培养基,其次为番茄燕麦、番茄煎汁、PSA、PDA和Richard培养基,而在2%水琼脂上明显慢于其他培养基(图2)。从菌落形态看,病菌在番茄燕麦、番茄煎汁、PSA、PDA和Richard培养基上菌落较为致密,在2%水琼脂和燕麦片培养基上菌落较为稀疏。从菌核产生数量看,在Richard培养基上产生最多,2%水琼脂培养基上产生最少。

图2 培养基对番茄茎基腐病菌菌丝生长的影响
2.3.1.2 碳源的影响
在淀粉和糊精培养基上,菌丝生长速度较快,其次为蔗糖、葡萄糖、甘露糖和果糖培养基,而在山梨糖培养基上明显较慢(图 3)。在葡萄糖、果糖、木糖、甘露糖、半乳糖、蔗糖、麦芽糖、乳糖及淀粉培养基上菌丝致密,无明显差异;在糊精培养基上,菌丝生长速度虽快,但菌丝稀疏,认为其并不适宜病菌菌丝生长。从菌核产生量看,除碳源山梨糖外,其他培养基上均能产生菌核,以碳源为淀粉和麦芽糖产生量最多,果糖和乳糖最少。
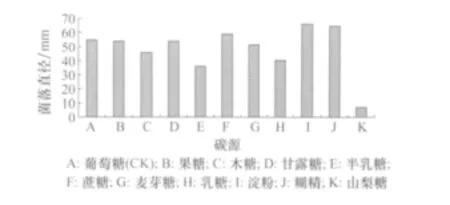
图3 碳源对番茄茎基腐病菌菌丝生长的影响
2.3.1.3 氮源的影响
测定结果见图4。

图4 氮源对番茄茎基腐病菌菌丝生长的影响
由图4结果表明,菌丝生长的适宜氮源为硝酸钾、硝酸钠、磷酸二氢铵、蛋白胨,以硝酸钠为最适,生长速度明显优于其他培养基。氮源为硫酸铵、氯化铵、精氨酸、胱氨酸、丙氨酸时生长较缓慢,在氮源为脲的条件下5d菌落未见扩展。除脲氮源外,菌丝在其他氮源条件下的生长均较为致密,无明显区别。从产生菌核数量看,氮源为蛋白胨时最多,而在硝酸钾、磷酸二氢铵、氯化铵、脲、精氨酸培养基上,20d后均未产生菌核。
2.3.1.4 温度的影响
测定结果表明,病菌菌丝在2~32℃温度范围内均可生长,适宜范围为20~25℃,最适为20℃。在2℃和32℃下生长极其缓慢(图5)。15~25℃下,病菌菌丝较为致密,其他温度下菌丝较为稀疏。考察菌核产生数量,以15℃产生量最多。
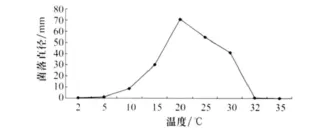
图5 温度对番茄茎基腐病菌菌丝生长的影响
2.3.1.5 pH的影响
病菌在pH3~10范围内均可生长(图6),但存在显著性差异。在pH为5~7时,菌落扩展最快,菌丝生长致密,为最适pH范围。病菌在pH为5~10范围内,菌丝生长均较为致密。从菌核产生数量看,pH7条件下产生最多,pH3条件下产生最少。
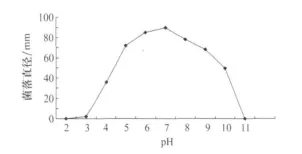
图6 pH对番茄茎基腐病菌菌丝生长的影响
2.3.1.6 光照的影响
图7测定结果可知,光照对菌丝生长没有明显作用,在全光照和半光照条件下,菌落直径相差很小;而光照和半光照条件比全黑暗条件下的菌落直径稍大一些,菌丝略致密一些,但均不明显。从菌核产生数量看亦无明显差异。

图7 光照对番茄茎基腐病菌菌丝生长的影响
2.3.2 菌丝致死温度
经≥50℃的几个温度处理后,菌丝均未生长,由此确定该菌菌丝致死温度为50℃、10min。
2.3.3 菌核萌发条件测定结果
2.3.3.1 碳源的影响
试验结果表明,不同碳源对菌核萌发的影响有明显差异,以麦芽糖为碳源时菌核的萌发率最高,3d萌发率可达100%,葡萄糖、蔗糖与甘露糖次之,3d萌发率达85%以上,而木糖的利用率最低,3d萌发率仅为60%。
2.3.3.2 温度的影响
不同温度下病菌菌核的萌发差异显著,在5~30℃范围内菌核均可萌发,20℃为最适温度,3d萌发率可达95%以上;而在0℃和35℃条件下不萌发。
2.3.3.3 pH的影响
菌核在pH3~10范围内均可萌发,适宜pH为5~7,以pH6的萌发率最高,3d萌发率可达98%;pH3菌核萌发率最低,3d萌发率仅为30%;pH2和pH11条件下菌核不萌发。
2.3.4 菌核致死温度
经≥59℃的几个温度处理后,菌核均不萌发,由此确定菌核致死温度为59℃、10min。
3 结论与讨论
研究结果表明,不同的营养、温度、pH等条件对番茄茎基腐病菌的菌丝生长和菌核产生有显著影响。菌丝在PDA培养基上、20℃、pH7的条件下生长最好;光照对菌丝生长和菌核产生没有明显的促进作用;多种碳源物质对菌核萌发均有一定促进作用,菌核在麦芽糖、20℃、pH6的条件下萌发率最高。
本研究结果表明,番茄茎基腐病的病原茄丝核菌(Rhizoctoniasolani)菌丝在2~32℃范围内均可生长,适宜范围20~25℃,最适20℃;低于2℃和高于32℃菌丝生长停止。结果与陈京元等人[9]记述的湿地松立枯丝核菌和赵桂东等人[10]记述的玉米、水稻和大豆立枯丝核菌,最高生长温度40℃的结果均有差异;而与赵桂东等人[10]记述的棉花立枯丝核菌和过崇俭[11]记述的水稻立枯丝核菌的最高生长温度30~35℃的研究结论相似。据此认为,病原同为茄丝核菌(Rhizoctoniasolani)的不同病害,由于其各自发生环境不同或是菌群分化等的影响,致使病原菌某些生物学特性也随之具有一定差异,尚待深入研究。
测定结果表明,病原菌菌丝的致死温度为50℃、10min,而人工培养的新鲜菌核的致死温度达到59℃、10min,其致死温度明显高于菌丝,说明菌核具有较强的抗逆性。生产实际中,该病菌抗逆性极强,病菌老熟菌核可在土壤中越冬存活几年而仍具有萌发和致病能力,生产上较难防治。本研究结果,对于生产上针对性拟定病害防治措施,具有一定指导意义。
[1]罗云生,刘永,张晓霞,等.保护地早春番茄茎基腐病的发生与综合防治[J].吉林蔬菜,2005(1):29.
[2]徐作珽,李林,李长松,等.番茄茎腐病病原鉴定及防治研究[J].植物病理学报,2004,34(3):286-288.
[3]向茂红,盛长存,贺天新.高台县番茄茎基腐病的发生与防治[J].甘肃农业科技,2007(5):48-49.
[4]陆家云.植物病原真菌学[M].北京:中国农业出版社,2002:60-363.
[5]孙广宁,宗兆锋.植物病理学实验技术[M].北京:中国农业出版社,2002:206-210.
[6]陆宁海,徐瑞富,吴利民,等.不同培养基对小麦纹枯病菌生长繁殖及致病力的影响[J].中国农学通报,2005,21(2):262-264.
[7]方中达.植病研究方法[M].北京:中国农业出版社,1998.
[8]周而勋,杨媚,李琳,等.培养基对水稻纹枯病菌菌丝生长和菌核形成的影响[J].华南农业大学学报,2002,7,23(3):33-35.
[9]陈京元,霍宪起,蔡三山,等.湿地松猝倒病病原鉴定及其生物学特性的观察[J].华中农业大学学报,2006,25(2):128-131.
[10]赵桂东,朱海波,周玉梅,等.大麦纹枯病菌与其轮作物丝核菌的生物学特性及致病力比较研究[J].大麦科学,1999(1):37-39.
[11]中国农业百科全书编辑部.中国农业百科全书:植物病理卷[M].北京:中国农业出版社,1996:438.
猜你喜欢
现代农业科技(2022年8期)2022-12-13
陕西农业科学(2022年2期)2022-03-28
麦类作物学报(2021年5期)2021-09-23
西北农业学报(2021年5期)2021-06-28
浙江农业学报(2021年5期)2021-05-31
中国食用菌(2021年1期)2021-03-12
农民致富之友(2020年16期)2020-06-19
南方农业·下旬(2019年6期)2019-10-30
中国食用菌(2019年9期)2019-10-21
新农业(2017年15期)2017-09-11
