
传染性胰腺坏死病毒VP3蛋白在干酪乳杆菌的表面表达
2010-08-06 08:04赵丽丽葛俊伟乔薪瑗刘巍巍赵永欣李一经
中国预防兽医学报 2010年1期
刘 敏,赵丽丽,葛俊伟,乔薪瑗,刘巍巍,赵永欣,李一经
(东北农业大学,黑龙江哈尔滨150030)
传染性胰腺坏死病病毒(Infectious pancreas necrosis virus,IPNV)是引起鲑科鱼类幼鱼肝胰腺等内脏实质器官出血、坏死为特征的病毒性传染病的病原。IPNV高致病力毒株对2月龄的幼鱼致死率高达90%,感染后幸存的鱼终生带毒,通过粪便和精卵排出病原[1-2],成为潜在的传染源。国内于1984年首次发现于西北地区某虹鳟养殖场,随后国内多家虹鳟养殖场先后暴发和流行。江育林等从患病的虹鳟(Oncorhynchus mykiss)体内分离到病原,并鉴定为IPNV sp血清型[3-6]。因该病流行范围广,发病和致死率高,给水产养殖业带来重大的经济损失。
IPNV属于双RNA病毒科(Birnaviridae),水生双RNA病毒属(Aquabirnavirus)的代表种。病毒粒子为无囊膜的二十面体,直径约60 nm[1]。基因组含有两个双链RNA节段被称作A段和B段,B段(2777 bp)编码一个较小的多肽VP1(94 ku),VP1是病毒粒子依赖的RNA聚合酶(RdRp),A段(3097 bp)内含一个大的开放阅读框架(ORF),编码由pVP2-VP4-VP3(106 ku)组成的多聚蛋白前体,在病毒成熟过程中,多聚蛋白前体再进一步裂解成VP2(54 ku)和VP3(31 ku)蛋白[2,7-8]。其中 VP2 和 VP3 是 IPNV 主要的结构蛋白,含有病毒主要抗原表位和中和抗原表位[2,9-10]。虽然大量研究已证明IPNV VP2蛋白是产生免疫保护的重要抗原,但是最新研究也表明,针对IPNV VP3蛋白的血清抗体,能够在细胞水平上抑制病毒对细胞病变效应(CPE),与VP2比较其免疫原性更强[11-12]。因此表明VP3蛋白亦含有重要的中和抗原表位。
目前对传染性胰腺坏死还没有特效治疗药物,主要以预防为主。欧州和美州地区的国家主要采取注射接种IPNV全病毒灭活疫苗预防传染性胰腺坏死的发生,然而仍每年有30%~40%的孵化场暴发传染性胰腺坏死,其中可能的原因是鲑鳟的幼鱼和成鱼在接种疫苗以前就感染了病毒[13]。理想的IPNV疫苗必须在稚鱼期发挥长效的保护,防止其成为病毒携带者。但是注射免疫接种不适用于稚鱼,因此口服和药浴是鱼类早期免疫接种的首选途径。国内目前对IPNV疫苗的相关研究还未见报道。
本实验将IPNV主要抗原VP3基因定向插入到乳酸杆菌表面表达型载体pPG1,以期获得IPNV VP3蛋白锚定在菌体表面,为有效刺激机体免疫应答产生免疫保护作用和潜在安全级重组活菌口服疫苗奠定重要的物质基础。
1 材料和方法
1.1 菌株与质粒 pMD18-T-VP3含有IPNV VP3(618 bp)基因序列,由本实验室构建保存;干酪乳杆菌表面表达载体pPG1,含有分泌信号肽基因序列(ssUSP)、氯霉素(Cm)抗性基因和锚定结构序列(anchor),Lactobacillus casei393由荷兰NIZO研究所惠赠;Lactobacillus casei393感受态细胞由本实验室制备保存。
1.2 主要试剂 抗兔HRP-IgG、抗兔FITC-IgG购自Sigma公司,用前分别用0.10 mol pH7.2 PBS液稀释成所需要的浓度;兔抗IPNV VP3蛋白高免血清由本实验室制备保存;质粒DNA小量提取试剂盒和柱式胶回收试剂盒购自上海华舜生物工程公司。
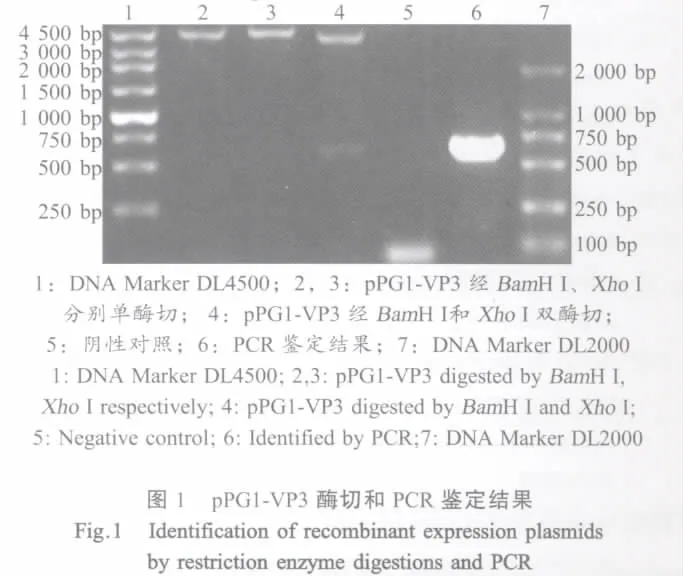
1.3 重组干酪乳杆菌表达载体的构建及鉴定 用质粒DNA小量提取试剂盒从大肠杆菌pMD18-T-VP3/JM109和含质粒pPG1干酪乳杆菌Lactobacillus casei 393中提取重组质粒pMD18-T-VP3及质粒pPG1。将pPG1和pMD18-T-VP3分别进行BamHⅠ、XhoⅠ双酶切后分别对目的片段用柱式胶回收试剂盒回收。将回收的pPG1和VP3的纯化产物进行连接,将连接产物电转化于感受态细胞Lactobacillus casei 393,取适量菌液涂布于含有10 μg/mL氯霉素(Cm)的MRS琼脂培养基上,37℃厌氧培养48 h~72 h,挑选菌落,提取质粒进行酶切和PCR鉴定阳性克隆,用于PCR鉴定的引物为P1(5'-AGCGGATCCT GCATCCGGGATGGACGAG-3')和P2(5'-AGCCTCGA GTCGTCGTTTCATCTGTC-3')。重组质粒命名为pPG1-VP3,含阳性重组质粒pPG1-VP3的干酪乳杆菌命名为pPG1-VP3/L.casei393。
1.4 目的蛋白的诱导表达 将pPG1-VP3/L.casei 393单个菌落接种5 mL MRS液体培养基中过夜培养使菌种活化,活化菌种按1∶20比例接种于10 mL含1%乳糖的MRS培养基中,30℃诱导16 h。
1.5 SDS-PAGE和western blot分析 分别取诱导前后样品各1 mL以12000 r/min离心5 min,菌体用100 μL 10 mg/mL的溶菌酶,37℃水浴作用60 min,12000 r/min离心5 min,弃上清后,加入1×PBS和等量的2×SDS-PAGE样品缓冲液(含DTT),混匀,沸水浴5 min,12000 r/min离心5 min。取上清,12%SDS-PAGE,并转印自硝酸纤维素膜上。兔抗IPNV VP3蛋白高免血清作为第一抗体,以辣根过氧化物酶标记羊抗兔IgG作为第二抗体,4-氯-1-萘酚底物显色溶液中显色15 min,观察结果。
1.6 间接免疫荧光检测 取经诱导的重组干酪乳杆菌和含空质粒的干酪乳杆菌各0.5 mL离心去上清后,分别用PBS洗菌体3次;加入兔抗IPNV VP3高免血清,混匀,37℃作用60 min,4000 r/min,离心5 min去上清,PBS离心洗菌体3次;加入稀释的羊抗兔IgG/FITC荧光抗体,悬浮混合后37℃作用60 min,4000 r/min离心5 min去上清,PBS离心洗菌体3次;菌体沉淀物悬浮于200 μL PBS缓冲液中,取适量涂片,自然干燥后,预冷丙酮固定30 min后,荧光显微镜观察。
2 结果
2.1 重组干酪乳杆菌表达质粒pPG1-VP3的鉴定重组质粒pPG1-VP3单酶切后得到大小约为4300 bp的基因片段,用BamH I、XhoI双酶切后得到大小为618 bp和3700 bp基因片段,以P1、P2为引物进行PCR扩增得到约为620 bp基因片段,与预计大小相符合(图1)。测序结果表明IPNV VP3基因片段已插入到表达载体pPG1中。
2.2 IPNV VP3蛋白在干酪乳杆菌中的表达与鉴定诱导后的菌体培养物经SDS-PAGE和western blot分析,结果表明在SDS-PAGE凝胶上未发现有明显的目的蛋白表达,但是转印NC膜上,经western blot检测在其位置(31 ku)出现明显的反应带(图2),说明目的蛋白在重组乳酸杆菌中获得了表达,并且表达蛋白能被抗血清所识别。以重组干酪乳杆菌pPG1-VP3/L.casei393和pPG1/L.casei393所进行的间接免疫荧光试验结果表明,诱导的重组干酪乳杆菌pPG1-VP3/L.casei393经荧光显微镜检查可见明显的黄绿色荧光(图3a),pPG1/L.casei393未出现荧光反应(图3b)。在间接免疫荧光试验中诱导后的重组菌体未经任何处理,由此表明IPNV VP3蛋白在重组干酪乳杆菌表面获得了表达。


3 讨论
一般认为VP3蛋白是作为IPNV的内衣壳蛋白参与病毒粒子的组成[14],但Tarrab等利用抗VP3单克隆抗体检测到IPNV病毒粒子表面存在部分VP3蛋白[15-16],Park等的研究表明,VP3蛋白含有重要的中和表位[11]。因此,由VP3基因编码的蛋白和VP2蛋白一样具有重要的免疫学功能。本实验利用具有锚定结构的乳酸杆菌表达载体,构建了表达IPNV VP3蛋白的重组干酪乳杆菌表达系统,SDSPAGE和western blot分析均表明,该蛋白获得了正确表达,间接免疫荧光检测表明,表达的重组蛋白锚定在菌体表面,从而为进一步研究该蛋白的免疫活性及其相关功能奠定了物质基础。为达到VP3蛋白表达的目的,在未影响表达蛋白抗原性的前提下,去除了疏水氨基酸碱基序列,使IPNV VP3蛋白在乳酸菌中得以表达。
Petter等利用大肠杆菌表达了IPNV VP2蛋白进行了免疫保护效果的测定,因表达蛋白在菌体内部以包涵体形式存在,需超声裂解后才能释放所表达的蛋白,而且大肠杆菌自身的毒性作用注射虹鳟会有一定的毒副作用[17-19]。Thomas等将VP2和VP3基因在酵母中进行了表达,表达后的蛋白需破碎酵母细胞壁后才能释放出来[20]。本研究采用具有锚定结构的乳酸菌表面表达载体,构建了重组干酪乳杆菌活菌载体系统,所表达的蛋白锚定于干酪乳杆菌菌体表面,不需要任何后处理,可直接作为鱼类的口服免疫制剂。利用活菌载体能够完整地将抗原信息传递到肠道黏膜免疫系统诱导免疫应答,同时乳酸菌本身就具有佐剂效应、黏附特性、非特异性抑菌抗菌,抗腹泻和肠道益生作用[21-22]。本项研究为防治鱼类感染IPNV的安全级乳酸菌重组活菌疫苗奠定了重要的物质基础。
[1]殷震,刘景华.动物病毒学[M].2版,北京:科学出版社,1997.
[2]Dobos P.The molecular biology of infectious pancreatic necrosis virus(IPNV)[J].Ann Rev Fish Dis,1995,5:25-54.
[3]牛鲁棋,赵志壮.东北地区虹鳟鱼IHN和IPN流行病学的初步研究[J].水产学报,1988,12(4):327-332.
[4]童裳亮,Heteick F M.山东虹鳟暴发传染性胰腺坏死病(IPN)[J].海洋通报,1989,18(1):116-117.
[5]江育林,徐伯亥,李伟,等.用酶联免疫吸附试验快速检测虹鳟的传染性胰腺坏死病毒[J].水生生物学报,1990,14(3):276-279.
[6]刘兴发,李红卫,钟志宏,等.虹鳟鱼传染性胰腺坏死病毒反转录聚合酶链反应的建立[J].中国兽医科技,1997,27(11):26-27.
[7]Duncan R,Mason C L,Nagy E,et al.Sequence analysis of infectious pancreatic necrosis virus genome Segment B and its encoded VP1 protein:a putative RNA-dependent RNA polymerase lacking the Gly-Asp-Aspmotif[J].Virology,1991,181:541-552.
[8]Gorbalenya A E,Pringle F M,Zeddam J L,et al.The palm subdomain-based active site is internally per mutedin viral RNA-dependent RNA polymerases of an ancient lineage[J].Mol Biol,2002,324:47-62.
[9]Frost P,Havarstein L S,Lygren B,et al.Mapping of neutralization epitopes on infectious pancreatic necrosis viruses[J].J Gen Virol,1995,76:1165-1172.
[10]Heppell J,Tarrab E,Lecomte J,et al.Strain variability and localization of important epitopes on the major structural protein(VP2)of infectious pancreatic necrosis virus[J].Virology,1995,214:40-49.
[11]Park J W,Jeong G.Identification of VP3 as an important neutralising epitope from DRT strain,a Korean isolate of infectious pancreatic necrosis virus(IPNV)[J].Fish and Shellfish Immunol,1996,6:207-219.
[12]Moon C H,Do J W,Park J W,et al.Comparison of the immunogenicity of recombinant VP2 and VP3 of infectious pancreatic necrosisvirus and marine birnavirus[J].Arch Virol,2004,149:2059-2068.
[13]Brun E,Evensen O,Rimstad E,et al.IPN in Salmonids:A Review.FHL&VESO[J].Trondheim,2003,51-67.
[14]Hjalmarsson A,Carlemalm E,Everitt E.Infectious pancreatic necrosis virus:identification of a VP3-containing ribonucleoprotein core structure and evidence for O-linked glycosylation of the capsid protein VP2[J].J Virol,1999,73:3484-3490.
[15]Caswell,Reno P,Nicholson B.Monoclonal antibodies to infectious pancreatic necrosis virus:analysis of viral epitopes and comparison ofdifferentisolates[J].JGen Virol,1986,67:2193-2205.
[16]Tarrab E,Berthiaume L,Heppell J,et al.Antigenic characterization of serogroup'A'of infectious pancreatic necrosis virus with three panels of monoclonal antibodies[J].J Gen Virol,1993,74:2025-2030.
[17]Petter F,Knut B,Curt E,et al.Analysis of the antibody response in Atlantic salmon against recombinant VP2 of infectious pancreatic necrosis virus(IPNV)[J].Fish and Shellfish Immunol,1998,8:447-456.
[18]Baneyx F.Recombinant protein expression inEscherichia coli[J].Curr Opin Biotechnol,1999,10:411-421.
[19]Marte B L,Suzanne B,Anthony E,et al.Antigenic comparison of a truncated form of VP2 of infec tious pancreatic necrosis(IPN)virus expressed in four different cell types[J].Fish and Shellfish Immunol,2001,11:203-216.
[20]Thomas A,Robert M,Christopher G,et al.Antigenicity of infectious pancreatic necrosis virus VP2 subviral particles expressed in yeast[J].Vaccine,2007,25:4880-4888.
[21]Perdigon G,Fuller R,Raya R.Lactic acidbacteria and their effect on the immune system[J].Curr Issues Intest Microbiol,2001,2(1):27-42.
[22]Tomohiko O,Yasuyuki A,Kenji Y,et al.Oral immunoadjuvant activity of a new symbioticLactobacillus caseisubspcaseiin conjunction with dextran in BALB/c mice[J].Nutrition Res,2005,(25):295-304.
猜你喜欢
今日畜牧兽医(2022年10期)2022-12-23
今日畜牧兽医(2022年10期)2022-12-23
当代水产(2022年1期)2022-04-26
食品界(2022年12期)2022-01-01
现代畜牧科技(2021年4期)2021-07-21
食品与发酵工业(2021年7期)2021-04-27
乳业科学与技术(2019年5期)2019-12-24
中国乳品工业(2018年7期)2018-08-21
中国调味品(2017年2期)2017-03-20
爱你(2016年20期)2016-11-25
