
鸡马立克病毒流行强毒株的致弱研究
2010-08-06 08:04刘爱玲刘长军张艳萍闫福海
中国预防兽医学报 2010年1期
刘爱玲,刘长军*,张艳萍,闫福海,2,张 锋
(1.中国农业科学院哈尔滨兽医研究所兽医生物技术国家重点实验室禽传染病研究室,哈尔滨150001;2.东北农业大学动物医学学院,哈尔滨150030)
马立克氏病(Marek's disease,MD)是由MD病毒(MDV)引起的鸡的一种高度接触性、传染性肿瘤病。依据血清学特性将MDV分为3个血清型,致瘤性MDV属于血清1型。MD可以通过疫苗免疫控制[1-2]。近年来,随着MDV毒力的增强,其致病力亦呈增强趋势,疫苗免疫失败的现象时有发生,因此,新的有效MD疫苗株的研究成为重点。由于与强毒株具有抗原相似性,血清1型的弱毒株将是更有效的疫苗。
我们从东北、四川2地区免疫发病鸡场中分离获得4株MDV流行毒株,选择在鸡胚成纤维细胞(CEF)上连续传代培养,获得4株高代次细胞毒株。我们对L-SYp85C和L-MSp75C毒株的体外生长特性、对无特定病原体(SPF)鸡的致病性及羽髓中复制能力等进行了动态的研究。本研究为了解传代毒株的弱化特性及弱毒疫苗株的筛选奠定了基础。
1 材料和方法
1.1 病毒株和鸡胚 分别命名为L-SY、L-MS、LCZ、L-ZY的MDV 4株流行毒株为本实验室从东北、四川2地区免疫发病鸡群采集病料分离获得(表1)。SPF雏鸭、SPF鸡胚、鸭胚由中国农业科学院哈尔滨兽医研究所实验动物中心提供。
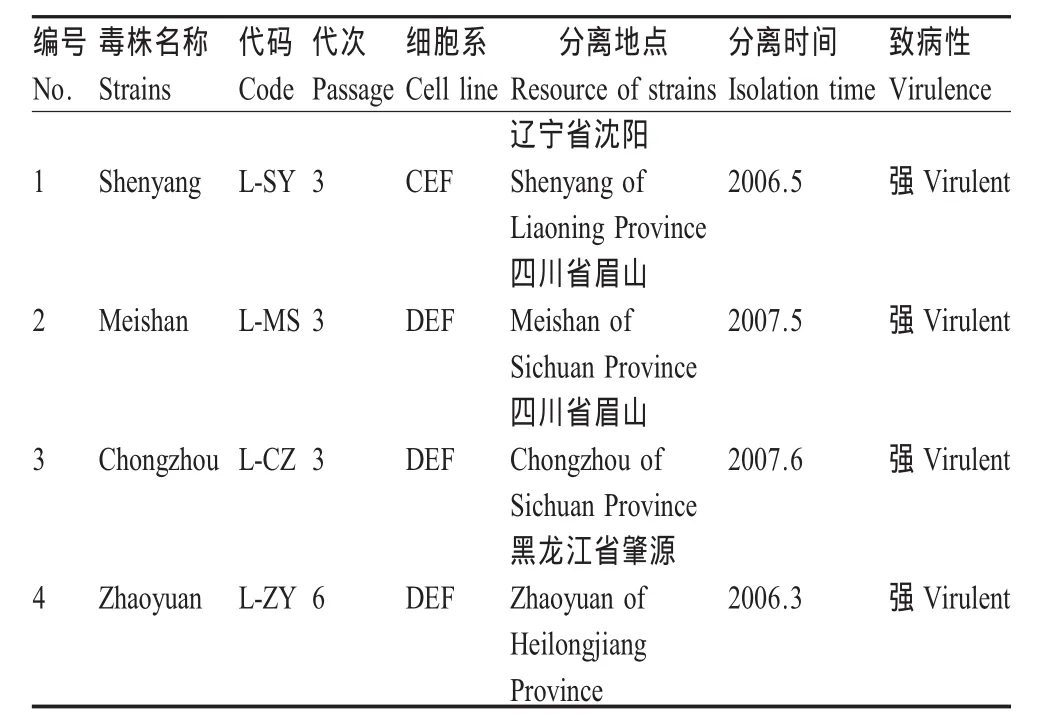
表1 实验用毒株Table 1 Strains used for the experiment
1.2 主要试剂 细胞培养用Medium199、犊牛血清购自美国GIBCO生物公司;PCR反应用试剂、Ex-Taq酶购自TaKaRa生物工程公司。
1.3 病毒连续传代培养 按常规方法用9日龄~10日龄SPF鸡胚制备CEF。将4株MDV分离毒株L-SY、L-MS、L-CZ、L-ZY分别接种于CEF,37℃CO2培养至出现细胞病变(CPE)时,胰酶消化,适量接种到新鲜的CEF中,如此反复传代培养至75代~85代,经克隆纯化,获得4株高代次细胞毒株。
1.4 体外生长曲线 通过测定相同接种剂量下不同生长时间的病毒滴度和每百万感染细胞中病毒基因组拷贝数2种方法比较病毒在传代过程中不同代次毒株增殖能力。选取L-SY毒株传代过程中第9代、30代、45代、65代及85代细胞冻存毒种,以同一批次细胞测定病毒滴度。将病毒原液做10倍递进稀释,以100 pfu剂量接种到单层CEF次代细胞的6孔培养板中,37℃CO2培养,在24 h、48 h、72 h、96 h、120 h、144 h时分别测定每个代次3个重复孔pfu数[3]。将病毒原液做10倍递进稀释,以100 pfu剂量接种已长成单层CEF次代细胞的6孔培养板中,37℃CO2培养箱中培养,在24 h、48 h、72 h、96 h、120 h、144 h分别收获每个代次病毒的3个重复孔,置于离心管中离心,PBS洗涤后再离心,消化,提取病毒基因组DNA,按照双重荧光定量PCR(FQ-PCR)方法[3]测定每百万感染细胞中的病毒基因组拷贝数。
1.5 132 bpr拷贝数检测 根据GenBank登录的GA株全基因序列(AF147806),采用Oligo6.0软件设计132 bpr基因引物,用于扩增4株分离毒株及其高代次细胞培养毒株、强毒J-1株和814疫苗株的132 bpr基因,引物由上海生物工程有限公司合成。上游引物序列:5'-TGCGATGAAAGTGCTATGG AGG-3',在GA株基因组的位置为127553~127574;下游引物序列:5'-GAGAATCCCTATGAGAAAGCG C-3';基因组中位置为 127848~127869。2拷贝PCR产物大小为314 bp,包含完整的132 bp重复序列。
PCR反应体积为50 μL。反应条件为:95℃5min;94℃30 s、56.5℃30 s、72℃45 s,25个~30个循环;PCR产物用2%的琼脂糖凝胶电泳检测,在BIO-RAD凝胶成像系统下观察电泳条带。
1.6 动物攻毒试验 将1日龄SPF雏鸡150只随机分成5个组,每组30只,负压隔离器中饲养至第12周。1组、2组鸡腹腔接种L-SYp85C毒株和L-MSp75C毒株,接种剂量为2×104pfu/只,3组、4组鸡腹腔接种L-SY毒株和L-MS毒株,接种剂量为2×103pfu/只,第5组鸡为空白对照组,腹腔接种稀释液。上述5组SPF鸡分别于接种后的7 d、14 d、22 d、29 d、45 d对每组试验鸡随机选取3只称重,剖检病理变化,采取MD典型病变组织做病理组织学观察,并采集3根~6根羽髓饱满的羽毛。
1.7 AGP试验 试验结束时,采集每组试验鸡羽髓丰富的羽毛,取羽髓、剪碎,加少许PBS混匀,作为被捡抗原样品,用MD阳性血清检测。
1.8 基因组的提取 细胞中病毒基因组的提取按常规方法进行。羽髓中DNA的提取方法:将1根~2根羽毛中的羽髓取出剪碎,加入裂解液500 μL,55℃消化4 h~6 h,酚氯仿抽提,无水乙醇沉淀DNA,溶于适量的TE缓冲液中,-20℃保存备用。
1.9 羽髓中病毒载量检测 羽髓中病毒载量的测定:按1.7中方法提取的样品DNA,通过测定同一样品管中FAM和ROX荧光强度,推导出样品DNA中MDV1基因组拷贝数和细胞拷贝数。数据采用MDV定量研究中常用的每百万细胞中病毒拷贝数(copies/106cell)的方法表示。
2 结果
2.1 分离毒株的传代培养 4株分离毒株经蚀斑纯化后,选用CEF连续传代至75代~85代,获得了4株高代次细胞毒株(表2)。L-SYp85C和L-MSp75C株在CEF适应性显著提高,表现为形成时间缩短,蚀斑面积增大,病变细胞间出现明显的融合现象。

表2 MDV毒株CEF传代Table 2 MDV strains passaged in CEF
2.2 体外生长曲线 L-SY毒株连续传代至85代,选取传代过程中第9代、30代、45代、65代及85代细胞冻存毒,通过测定不同生长时间的病毒滴度及每百万感染细胞中病毒基因组拷贝数2种方法绘制病毒的体外生长曲线。这2种检测方法检测对象有差异,生长曲线也不同。对生长过程中病毒滴度的检测是对具有感染性的活病毒细胞进行的检测,可反映病毒的感染能力;而对每百万感染细胞中病毒基因组拷贝数的检测则反映了全部的被感染细胞,而与病毒感染力不相关。2种方法绘制的L-SY毒株传代过程中生长曲线均表明,随细胞传代代次递增生长速度显著增加(图1,图2)。
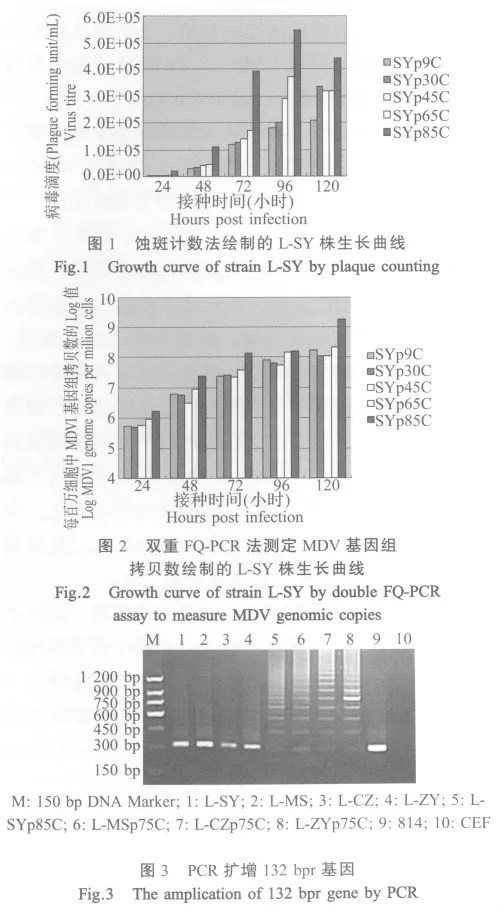
2.3 132 bpr基因拷贝数变异 132 bpr基因拷贝数增加被认为是毒株弱化的一个特征性的变化[12]。132 bpr基因在毒力较强的毒株中一般以2拷贝或3拷贝的形式存在。4株MDV分离毒株的132 bpr基因经PCR扩增鉴定为2个拷贝,在CEF中连续传代培养后获得了4株高代次细胞毒株,其132 bpr基因拷贝数发生变异,凝胶电泳图可见由2~7拷贝等形成的多拷贝混合体,最多可达到10个拷贝以上(图 3)。
2.4 致病性试验 试验结束时将所有鸡迫杀、剖检,与对照鸡比较分析。L-SY、L-MS毒株攻毒鸡出现典型MD临床症状,鸡体瘦小,精神沉郁,羽毛粗乱等,并伴有早期的死亡。剖检主要表现有不同程度的腺胃肿瘤、性腺肿瘤、肺脏肿瘤、心脏肿瘤等,灰白色,质地坚硬而紧密。L-SYp85C毒株攻毒鸡剖检仅有部分鸡表现为性腺不同程度肿大和盲肠扁桃体出血等炎症反应。

表3 L-SYp85C、L-MSp75C毒株致病性试验Table 3 The experiment for pathogenicity of strain L-SYp85C and L-MSp75C
L-MSp75C毒株试验组和L-SYp75C毒株试验组体重平均值分别对照组比较分析,2组间总体平均数差异不显著;但是L-MS、L-SY毒株试验组与对照组比较,差异显著(p<0.05)(表3)。
AGP试验检测L-SY、L-MS毒株攻毒鸡羽髓抗原,MD阳性率为100%;L-MSp75C毒株和L-SYp75C毒株攻毒鸡羽髓中AGP抗原检测均为MD阴性。
2.5 病理组织学变化 L-SY和L-MS毒株攻毒鸡病理组织学变化主要表现为卵巢内卵泡萎缩,部分卵泡消失,间质中伴有大面积弥散性淋巴细胞浸润及曲细精管严重坏死崩解;腺胃粘膜上皮有脱落,腺管内有淋巴细胞浸润灶,固有层内有大量淋巴细胞浸润;法氏囊有部分滤泡内细胞全部消失,滤泡呈大空泡状,有的滤泡中心出现嗜中性粒细胞浸润灶等。L-SYp85C、L-MSp75C毒株感染的鸡只大部分同空白对照组鸡只相似,无病理组织学变化;少数鸡表现为性腺曲细精管萎缩和部分坏死,盲肠扁桃体淋巴小结细胞内的轻度减少和出血。
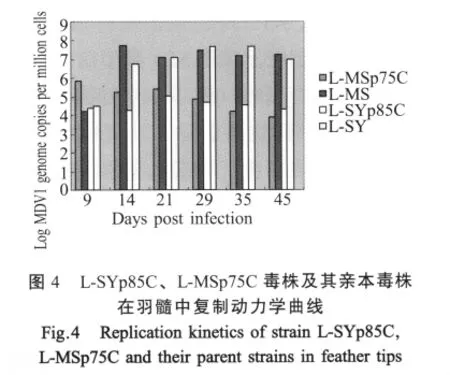
2.6 羽髓中病毒复制能力 通过检测发现,高代次细胞毒株L-SYp85C和L-MSp75C在整个检测期间病毒载量均低于106copies/106cells,并且在接种21 d后病毒载量开始下降,而L-SY和L-MS毒株感染14 d后,羽髓中病毒载量分别为106.8copies/106cells和 107.7copies/106cells,均在 106copies/106cells以上,并在14 d~21 d基本保持同一平台。这说明这2株传代毒株在鸡体内的复制能力显著降低。而2株传代毒株在检测的7 d~21 d病毒载量相对升高,推测是由高剂量病毒接种导致的结果。
3 讨论
将MDV通过细胞培养连续传代培育致弱疫苗已成功地应用了40年。属于血清Ⅰ型的HPRS-16att是第一株商品化的MDV疫苗,它是在鸡肾细胞上传代33代而致弱的。CVI 988是在鸭胚培养26代~35代致弱的。研究表明,MDV强毒株在细胞中连续传代致弱所需要的代次既取决于分离毒株的毒力,也与继代策略有关。GA/22、596A、617A等强毒株完全致弱需CEF继代70代~80代,645、660A、686等超强毒株则需80代~120代才能达到完全致弱。MDV流行强毒株通过细胞连续传代致弱获得了许多疫苗候选毒株,如Md11/75或R2/23(Md11p106)等。
本实验选用的4株MD分离毒株是近年来本实验室从东北、四川2地区免疫发病鸡群中采集病料分离获得,其中L-SY、L-ZY株动物实验及L-SY、L-MS、L-CZ毒株羽髓中复制能力研究表明这4株毒株为流行强毒株[4-5]。4株分离毒株采用CEF连续传代培养获得了4株高代次细胞毒株。分离毒株体外连续传代培养后,病毒生物学特性发生明显变化,表现为生长速度加快,病毒滴数提高。
病毒在细胞传代培养后生长速度的增加往往导致其在鸡体内的复制能力降低。我们的研究也证实传代毒株具有体内复制能力弱的特点。Baigent等通过实验证明了羽髓中病毒载量的检测是比较理想检测材料,羽髓中病毒载量反映了病毒体内的复制能力和毒株毒力[6-7]。Reddy和Yunis等通过定量感染鸡的MDV1载量,得出MDV1早期复制率高其毒力强,MD发病率、死亡率高的结论[8-9]。我们将传代获得的分离毒株L-SYp85C、L-MSp75C毒株以亲本毒株10倍剂量人工感染SPF鸡,检测其在7 d~45 d羽髓中的病毒载量发现,L-SYp85C、L-MSp75C毒株羽髓中病毒载量在整个检测期内均明显低于亲本毒株羽髓中病毒载量,既高代次毒株在羽髓中复制能力降低。而且病理组织学观察和体重均值的统计学分析也表明,这2株毒株经传代培养后毒力降低。弱毒疫苗株在鸡体内的复制能力与其对强毒攻击的免疫保护能力密切相关。Gimeno等将一组具有高度免疫力的候选疫苗株与另一组保护力较差的候选疫苗株进行了比较,他们发现,有高度保护力的疫苗株往往在体内的复制能力较强,而且,能诱发鸡的辅助性和细胞毒细胞的增殖[10-11]。因此,对MDV强、弱毒株羽髓中病毒载量检测有助于了解病毒的体内复制情况,判定毒株毒力,进而为疫苗的开发奠定基础。
此外,检测132 bpr基因的拷贝数常作为判定毒株毒力的重要指标。Becker等对不同毒力毒株132 bp重复区的鉴定说明弱毒株通常具有CVI988类似的132 bp序列[12]。我们采用PCR方法扩增4株高代次细胞毒株及其亲本毒株、J-1株和814疫苗株的132 bpr基因,4株亲本毒株132 bp重复区PCR鉴定均为2个拷贝,但传代培养获得的高代次细胞毒株其拷贝数明显增加,可达到10个拷贝以上,符合弱毒株的特点。
[1]Saif Y M.苏敬良译.禽病学[M].北京:中国农业出版社,2005.
[2]殷震,刘景华.动物病毒学[M].北京:中国科学出版社,1997.
[3]张颖,刘长军,秦运安,等.应用双重荧光定量PCR方法检测马立克氏病毒血清1型病毒[J].中国预防兽医学报,2007,29(1):46-51.
[4]施维松,刘长军,张艳萍,等.4株鸡马立克氏病病毒国内分离株Meq基因的克隆与序列分析[J].病毒学报,2008,2:128-125.
[5]李晶梅,刘长军,张艳萍,等.鸡马立克氏病病毒在鸡淋巴细胞和羽髓中的复制动力学分析[J].中国预防兽医学报,2008,30:880-884.
[6]Baigent S J,Smith L P,Currie R L.et al.Replication kinetics of Marek's disease vaccine virus in feathers and lymphoid tissues using PCR and virus isolation [J].J Gen Virol,2005,86:2989-2998.
[7]Baigent S J,Smith L P,Nair V K,et al.Vaccinal control of Marek's disease:current challenge,and future strategies to maximize protection[J].Vet Immunol Immunopathol,2006,112:78-86.
[8]Reddy S M,Witter R L,Gimeno I.Development of a quantitative competitive polymerase chain reaction assay for serotype 1 Marek's disease virus[J].Avian Dis,2000,44,770-775.
[9]Yunis R,Jarosinski K W,Schat K A.Association between rate of viral genome replication and virulence of Marek's disease herpesvirus strains[J].Virology,2004,328:142-150.
[10]Gimeno I M,Witter R L,Hunt H D.et al.Biocharacteristics shared by highly protective vaccines against Marek's disease[J].Avian Pathol,2004,33(1):59-68.
[11]Witter R L,Calnek B W,Buscaglia C,et al.Classification of Marek's disease viruses according to pathotype-philosophy and methodology[J].Avian Pathol,2005,34:75-90.
[12]Becker Y,Tabor E,Asher Y,et al.PCR detection of amplified 132 bp repeats in Marek's disease virus type 1(MDV-1)DNA can serve as an indicator for critical genomic rearrangement leading to the attenuation of virus virulence[J].Virus Genes,1993,7(3):277-287.
猜你喜欢
林业与环境科学(2022年2期)2022-07-12
中国医学工程(2022年5期)2022-05-27
右江民族医学院学报(2022年2期)2022-05-19
中国乳品工业(2022年4期)2022-05-19
河北医学(2021年10期)2021-10-27
昆明医科大学学报(2020年11期)2020-12-28
中西医结合肝病杂志(2020年2期)2020-10-27
中国临床医学影像杂志(2019年6期)2019-08-27
科学与财富(2016年32期)2017-03-04
中华实验和临床病毒学杂志(2016年3期)2016-08-09
