
陆地棉与海岛棉抗氧保护酶差异的比较
2010-09-20 00:24胡根海张金宝朱颜平刘向梅王清连
东北农业大学学报 2010年4期
胡根海,张金宝,朱颜平,刘向梅,王清连
(河南科技学院生命科技学院,河南 新乡 453003)
植物体生长发育所需要的物质和能量主要通过有氧代谢提供。在有氧代谢过程中,不可避免的产生一定量自由基,如超氧阴离子、羟基自由基、过氧化氢等。当植物处于极端温湿度、干旱、盐胁迫、除草剂、二氧化硫和辐射等逆境时,也会产生大量活性氧,大量自由基和活性氧的存在会给植物体造成严重的损伤。SOD、POD和CAT是生物体内重要的自由基清除剂,在清除机体内多余的超氧阴离子、解除超氧阴离子对生物体的毒性过程中起着关键性作用[1-2]。海岛棉和陆地棉是生产上两大栽培种,海岛棉表现生长旺盛植株高大,叶色暗绿,优质长绒,但是产量低。与其相反,陆地棉植株矮小,叶小色淡,绒长(陆地棉21~33 mm)较短(与海岛棉33~39 mm比较),但产量高。利用陆海杂交改良陆地棉是育种家最早选择的方法,但陆海杂交后代分离严重,目标性状很难整合在一起,探讨陆海之间抗氧保护酶动态变化的差异,找到各指标的最佳检测时期,有可能为利用陆海杂交培育新品种的杂交后代选育提供参照数据。目前对同一种植条件下陆地棉与海岛棉抗氧保护酶活性变化的差异性研究尚未见报道。本研究旨在探讨陆地棉与海岛棉在相同栽培条件下,抗氧保护酶活性变化的动态变化差异性,为黄河流域棉区利用海岛棉改良陆地棉提供理论上的参考依据。
1 材料与方法
1.1 材料
试验在河南科技学院棉花实验基地进行。选用7个陆地棉和4个海岛棉品种(系)作供试材料(部分材料购自中国农业科学院棉花研究所品种资源室),供试棉花材料名称:中棉所50、中棉所64、早熟0式、旱零1号、鸡脚叶棉、新疆01、新疆02、新海18、跃零1号、跃零2号和海岛棉1号。棉花材料于2008年5月20日大田播种,按照常规大田管理方式管理,在棉花发育中的6月18日(5片真叶苗期),7月4日(现蕾期),7月20日(始花期),8月6日(花铃期)和8月26日(吐絮期)取样,样品取自同期棉花相同部位的全展功能叶片。
1.2 方法
称取剪碎的新鲜棉花叶片共3份,每份0.3 g左右,放入试管中,测定时至少重复3次,取平均值。超氧化物歧化酶(SOD)活性测定采用硝基四氮唑蓝光化还原法加以改进[3],过氧化氢酶(CAT)活性测定采用紫外吸收法[3],过氧化物酶(POD)活性测定采用愈创木酚显色法,以每分钟内A470变化0.01为1个酶活性单位[3]。所测数据均采用Excel软件分析。
2 结果与分析
2.1 海岛棉与陆地棉叶片的SOD活性变化差异
由图1可知,陆地棉和海岛棉的SOD活性变化在整个生育期中呈规律性变化,陆地棉和海岛棉的变化趋势有差异。陆地棉总体表现从6月18日(苗期)开始下降,7月4日(现蕾初期)至7月20日(始花期)保持稳定,7月20日以后快速升高;4份海岛棉表现在整个生育期中SOD活性表现出波动上升的基本趋势。6月18日至7月4日基本维持恒定,7月4日后上升,在7月20日出现高峰,然后下降,8月6日后上升,从整体上看,所有材料均是8月26日吐絮期最高。
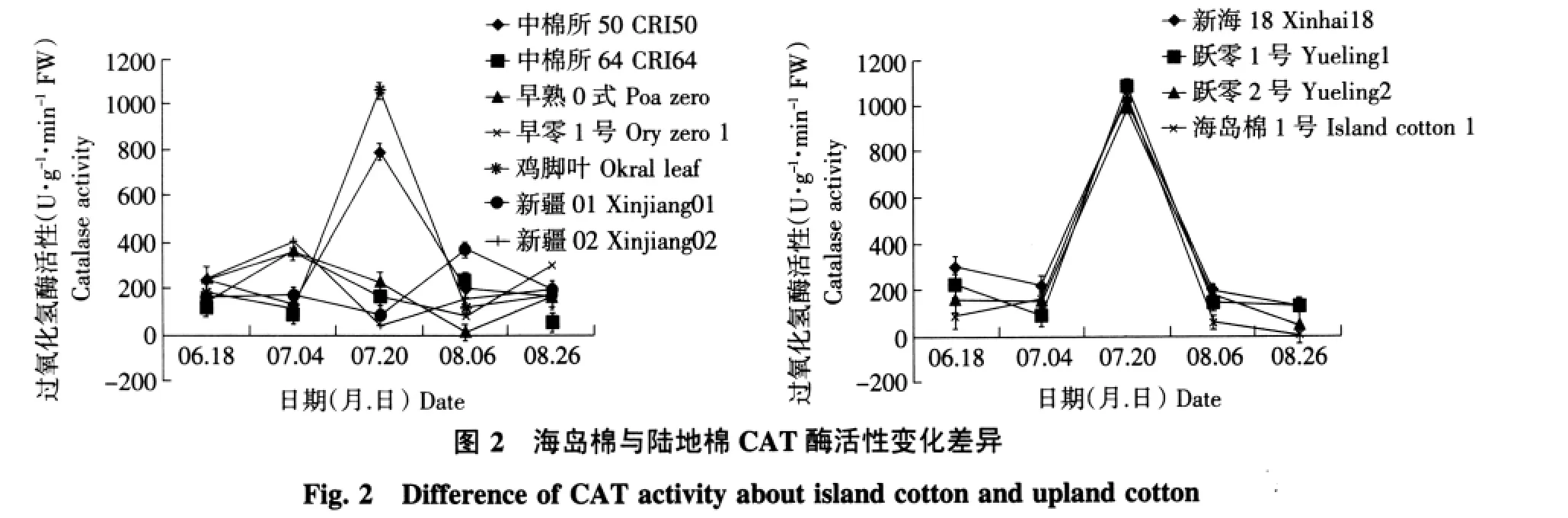
2.2 海岛棉与陆地棉叶片的CAT活性变化差异
由图2可知,陆地棉7份材料中不同材料之间CAT活性有不同变化规律,总体看每个材料都有单一峰值,但不同材料峰值出现时期,最高值(极值)不同;新疆02、鸡脚叶棉和早熟0式峰值出现较早在7月4日(现蕾初期),中棉所50和旱零1号峰值出现在7月20日(始花期),中棉所64、新疆01峰值出现在8月6日(花铃期);海岛棉在整个生育期中CAT活性与陆地棉相似也表现出单峰曲线,但4份海岛棉变化一致,6月18日至7月4日基本维持恒定或略下降,7月4日后上升,在7月20日出现高峰,然后下降,8月6日后维持较低的水平。所有材料均是7月20日(始花期)最高,而且不同材料间酶活性差异较小。
2.3 海岛棉与陆地棉叶片的POD活性变化差异
由图3可知,在整个生育期中,陆地棉和海岛棉的POD活性变化趋势非常相似,仅在6月18日至7月4日略有差异。7份陆地棉均表现6月18日至7月4日略微下降,从7月4日后上升,后期不同材料上升幅度有差异。海岛棉POD活性表现出整体前期较低后期较高的上升趋势,所有材料均是8月26日吐絮期最高。不同材料间差异表现为:跃零1号与陆地棉相似,从6月18日至7月4日略下降;7月4日后所有材料都是上升趋势,在8月26日新海18与其他3个材料有较大差异。


3 讨论与结论
生物体的一切生命过程实际上都是一系列酶促反应的生化过程,酶含量与酶活性变化往往意味着该酶专一性催化的生化过程变化,反映了生物体某一特性的变化。在自由基伤害学说中,SOD、CAT和POD是防御系统的主要保护酶类,这三者协调一致,使生物体内自由基维持在一个较低水平,从而防止自由基伤害。宋美珍等在短季棉抗氧保护酶遗传研究中认为SOD、CAT和POD存在加性效应,并找到其相应的QTL[2,4-7],为短季棉抗氧保护酶育种利用提供依据。本试验结果表明,海岛棉和陆地棉一样,其抗氧保护酶在整个生育期中也存在变化规律,但陆海之间存在差异,这为陆海杂交后代选育提供重组的可能。
从海岛棉与陆地棉的SOD活性变化比较可以看出,陆地棉与海岛棉有不同的变化规律,试验得到的陆地棉动态变化趋势与前人结果基本一致[7],试验结果差异主要在花铃期以后,宋美珍等认为花铃期以后为下降趋势[7]。本试验得到上升趋势,作者认为花铃期以后气温逐渐降低,有可能是外界的低温诱导激活了棉株的SOD合成机制,导致后期较高的SOD酶活性,后期较高的酶活性可以清除低温引起自由基的增加更有利于棉花生长。陆地棉和海岛棉都是后期SOD酶活最高。已有研究结果认为,大豆SOD活性在整个生育期中呈现规律性变化,并存在品种间的差异[8-9]。梁洪艳等研究发现,小麦品种SOD活性与其抗盐碱紧密相关[10]。结合前人结论,试验结果建议,在整个生育期中,陆地棉、海岛棉的SOD活性与其他作物一样具有规律性;陆地棉SOD高活性期在花铃期以后,筛选高SOD活性材料应在此期比较适宜;海岛棉SOD高活性期在始花期或花铃期以后,建议可以选择在花铃期以后测定评价SOD。
从海岛棉与陆地棉的CAT活性变化比较可见,陆地棉不同材料有不同的变化规律,郭明等研究结果证实农药喷施可以引起棉花CAT的涨落[11],李永平等发现大豆CAT活性具有明显的基因型差异[12],姜佰文等认为在大豆四个生长阶段中,CAT活性的变化规律呈单峰曲线,初花期最高[13]。本试验是统一管理,材料种植在一个小区内,试验数据有可比性,结合其他作物认为棉花的CAT在整个生育期中是有规律可循的,但陆地棉不同材料不同变化可能是其诱导反应机制不同,海岛棉有较好的规律性。根据图2建议评价CAT的最佳时期,海岛棉为始花期,陆地棉需先预测其最佳评价时期再做决定。
从海岛棉与陆地棉的POD活性比较可见,海岛棉和陆地棉的整个生育期都是前期低,后期较高,李付广等研究发现耐盐品种的子叶POD活性明显高于不耐盐品种[14],丁伟等研究豆磺隆胁迫对甜菜质膜造成的伤害时发现,随着施药后时间的延长,叶片中POD、SOD活性均呈增加趋势[15]。 依试验结果结合其他作物资料可知:在整个生育期中,海岛棉、陆地棉都存在POD酶活性较高的材料,在育种新材料选育时,可以从增加抗逆性的角度,通过测定材料POD酶活性,比较其高低,选择出高POD活性的新材料。根据图3变化趋势,评价POD活性可能在吐絮期较好。
[1]方允中,郑荣粱.自由基生命科学进展:第3集[M].北京:原子能出版社,1993.
[2]喻树迅,宋美珍,范术丽,等.短季棉早熟不早衰生化辅助育种技术研究[J].中国农业科学,2005,38(4):664-670.
[3]张志良,瞿伟菁.植物生理学实验指导[M].3版.北京:高等教育出版社,2003.
[4]Yu S X,Song M Z,Fan S L.Biochemical genetics of short-season cotton cultivars that express early maturity without senescence[J].Journal of Integrative Plant Biology Formerly Acta Botanica Sinica,2005,47(3):334-342.
[5]宋美珍,喻树迅,范术丽,等.短季棉主要农艺性状的遗传分析[J].棉花学报,2005,17(2):94-98.
[6]宋美珍,喻树迅,范术丽,等.短季棉早熟不早衰生化性状的遗传分析[J].西北植物学报,2005,25(5):903-910.
[7]宋美珍,喻树迅,范术丽,等.早熟不早衰短季棉品种(系)及其杂交后代抗氧化酶活性的变化[J].棉花学报,2006,18(1):63-64.
[8]赵燕丽,于龙凤,李茫雪,等.大豆叶片和豆荚与衰老相关的某些生理特性比较[J].东北农业大学学报,2008,39(1):6-9.
[9]钟子楠,李淑芹,闫爱博,等.碱石灰改良酸土对大豆植株保护系统的影响[J].东北农业大学学报,2009,40(1):46-48.
[10]梁洪艳,马凤鸣,杨德光,等.Na2CO3胁迫对春小麦苗期生理特性的影响[J].东北农业大学学报,2008,39(1):19-22.
[11]郭明,徐雅丽,刘明,等.几种农药对棉花过氧化氢酶过氧化物酶活性的影响[J].农业环境保护,2001,20(1):l0-12,22.
[12]李永平,王长发,赵丽,等.不同基因型大豆叶片生理特性的研究[J].干旱地区农业研究,2009,27(1):173-177.
[13]姜佰文,戴建军,王春宏,等.干旱胁迫下硼对大豆植株保护酶活性的影响[J].作物杂志,2009(1):50-53.
[14]李付广,李凤莲,李秀兰.盐胁迫对棉花幼苗保护酶系统活性的影响[J].河北农业大学学报,1994,17(3):52-55.
[15]丁伟,陶波,程茁,等.甜菜对豆磺隆敏感性与质膜过氧化关系的研究[J].东北农业大学学报,2005,36(5):556-560.
猜你喜欢
中国土壤与肥料(2021年5期)2021-12-02
江苏农业科学(2017年22期)2018-01-06
江苏农业科学(2016年5期)2016-07-23
西南农业学报(2016年5期)2016-05-17
中国种业(2016年3期)2016-04-14
西藏科技(2015年5期)2015-09-26
山东农业科学(2014年1期)2015-03-09
新疆农垦科技(2014年10期)2014-02-28
新疆农垦科技(2014年5期)2014-02-28
植物营养与肥料学报(2011年2期)2011-10-26
