
饲粮蛋白质水平对肥育猪肌肉嫩度及背最长肌钙调磷酸酶-活化T细胞核因子信号途径的影响
2011-03-14 05:33朱宇旌邵彩梅
动物营养学报 2011年7期
张 勇 陶 亮 崔 岩 朱宇旌 邓 科 孙 璀 邵彩梅
(1.沈阳农业大学畜牧兽医学院,沈阳 110866;2.辽宁禾丰牧业股份有限公司,沈阳 110164)
保证肉品质是保障养殖业、肉类加工业、批发零售业效益及广大消费者的切身利益的重要因素,对肉品质进行营养调控是当今动物营养领域研究的热点之一。钙调磷酸酶(calcineurin,CaN)、钙调蛋白(calmodulin,CaM)和活化T细胞核因子(nuclear factor of activated T cells,NFAT)共同构成了钙调磷酸酶-活化T细胞核因子(CaNNFAT)信号途径,这一途径在调节骨骼肌生长,决定肌纤维类型特征中起着重要作用[1]。钙蛋白酶抑制蛋白(calpastatin,CAST)是内源性的,需要Ca2+信号来活化。它可以抑制肌肉内蛋白质降解,降低肌细胞生长速度;动物被屠宰后,它可抑制钙蛋白酶的活性,降低蛋白质水解[2]。CaN是受Ca2+信号活化的一种多功能信号酶,激活的CaN可以使NFAT的多个丝氨酸残基去磷酸化,从而导致NFAT的构象发生变化,促使NFAT在细胞核内定位并使DNA结合功能域暴露,转录因子NFAT脱磷酸而活化而进入核内[3]。近年来,大部分的研究都集中在Ca2+-CaN-NFAT信号途径在骨骼肌生长、发育及功能维持中的生物学功能[4-8],有关 CAST作为肉质候选基因在研究牛、猪等哺乳动物中有所报道[9-12],而饲粮蛋白质水平对CAST和CaN-NFAT信号途径相关蛋白mRNA表达量的影响以及CaN-NFAT信号途径与肌肉嫩度的关系都鲜有报道。本研究利用实时定量PCR(real time-PCR,RT-PCR)方法,以β-肌动蛋白(β-actin)为内参,研究饲粮中不同蛋白质水平对CAST、CaN、NFAT和CaM mRNA在背最长肌相对表达量的影响,明确CAST与CaN-NFAT信号途径在猪肌肉嫩度调控中的作用,为改善猪肉品质提供科学依据。
1 材料与方法
1.1 试验仪器
数显式肌肉嫩度仪(C-LM 3型,东北农业大学工程学院研制);高速台式冷冻离心机(TGL-20M,长沙湘仪离心机仪器有限公司);紫外分光光度计(Cary 50 Probe,美国Varian公司);基因扩增仪[TC-96/G/H(6),杭州博日科技有限公司];PCR仪(ABIPRISM 7000 HT Real-Time RCRSystem,美国ABI公司);凝胶成像系统(江苏捷达)。
1.2 试验试剂
RNA提取试剂盒(RNAiso Reagent)、RT-PCR试剂盒[SYBRPrimeScriptTMRT-PCR Kit(Perfect Real Time)]、DEPC 水、DNA 分子量标准(DL2000)、50 bp DNA Ladder Marker均购自于TaKaRa公司;琼脂糖购自于宝泰克(西班牙)公司;其他常用化学试剂均为国产分析纯。
1.3 试验动物与试验设计
本试验采用完全随机区组试验设计,选择初始重约50 kg的杜洛克×长白×大白三元杂交猪90头,随机分配到3个处理,每个处理3个重复,每个重复10头,公母各占1/2。3个处理分别采用12%、16%、20%理想蛋白质水平的饲粮,饲粮能量水平相同,正试期58 d,参照NRC(1998)肥育猪的营养需要配合成粉状全价料,饲粮消化能保持相同。饲粮组成及营养水平见表1。
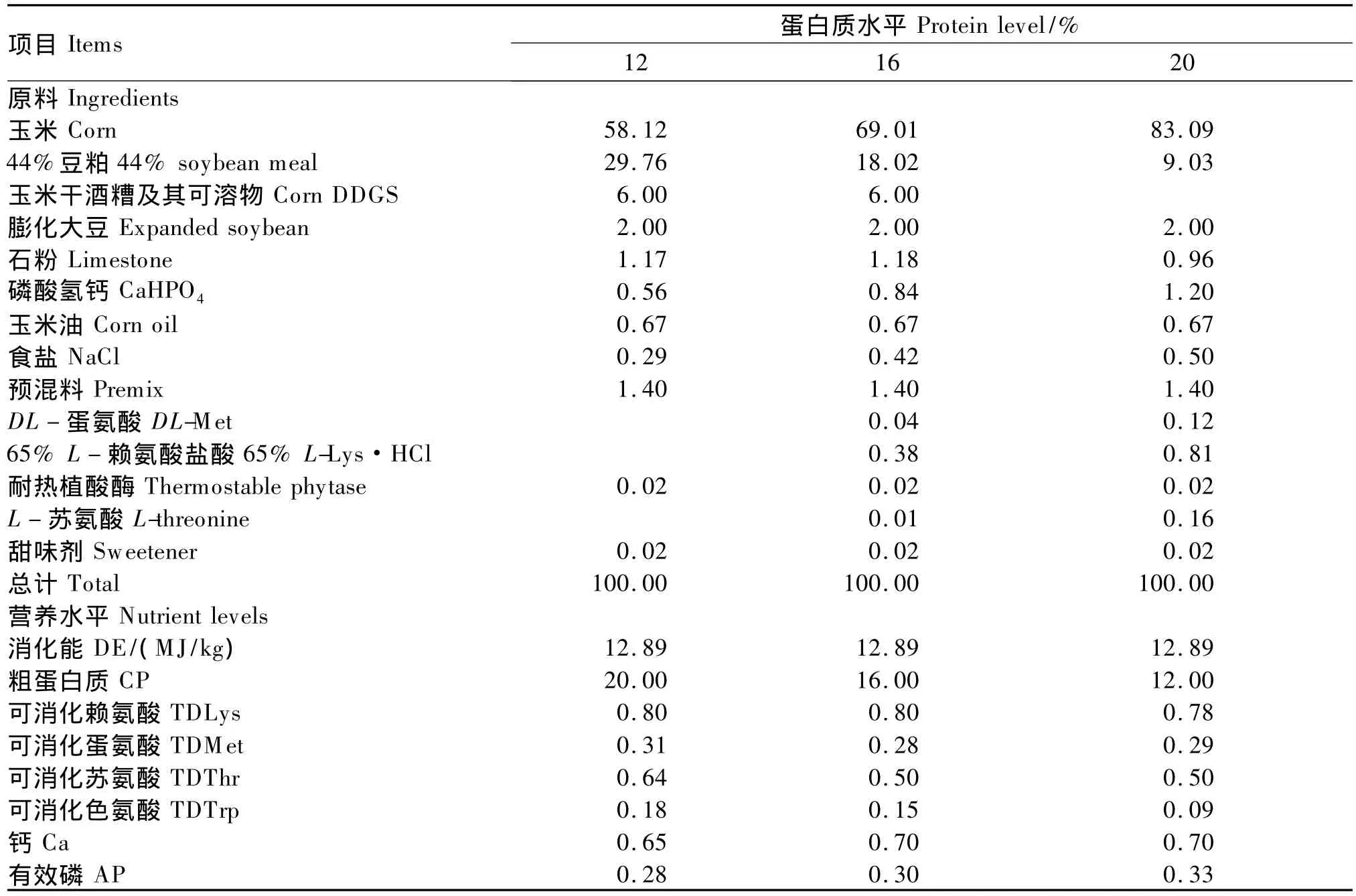
表1 饲粮组成及营养水平(风干基础)Table 1 Composition and nutrient levels of diets(air-dry basis) %
1.4 试验设计与饲养管理
试验前进行驱虫、防疫、编号等工作。试验猪水泥地面单圈饲养,自由采食,以鸭嘴式饮水器提供充足清洁饮水,日饲喂3次,清粪3次,保持舍内清洁,随时观察并记录饲养期间猪的食欲、精神状况、粪便等情况。
1.5 样品采集
为了使胴体尽量少受破坏,肉样采集位置为猪最后肋骨和最后腰椎间的单侧背最长肌,采样400 g,用于肉品质测定;并在最短的时间内于第13~16肋骨间取猪背最长肌样本30 g,放入锡箔纸中包好,用液氮迅速冷冻,之后放于-80℃保存备用。
1.6 肌肉嫩度的测定
采用C-LM 3型数显式肌肉嫩度仪测定肉样剪切力,具体试验方法参考张勇等[13-14]文献。
1.7 RT-PCR法测定相关蛋白m RNA表达量
1.7.1 总 RNA 提取
参照RNA提取试剂盒推荐方法,提取猪背最长肌样品总RNA。提取后的RNA用紫外分光光度计在260和280 nm处测定吸光度值,根据OD260nm/OD280nm检验纯度,并按如下公式计算RNA的浓度:

1.7.2 cDNA 合成
按照RT-PCR试剂盒操作方法进行。cDNA合成后放-20℃保存备用。
1.7.3 引物设计与合成
β-actin、CaN、NFAT、CaM 和 CAST 的引物序列及参数见表2。引物均由上海生工有限公司合成。
1.7.4 RT-PCR
参照试剂盒操作说明,采用SYBR Green荧光染料法,在RT-PCR仪上进行相对定量分析。反应条件:94℃变性30 s,58~62℃退火30 s(具体基因的退火温度见表2),72℃延伸45 s,40个循环。

表2 引物序列及实时定量PCR反应条件Table 2 Primer sequences and the conditions for real-time PCR
1.7.5 相对表达量计算
目的基因相对表达量按以下公式计算:

目的基因相对表达量=2-△△Ct。
式中:Ct为阈值循环。
1.8 数据统计与分析
用中国科学院沈阳应用生态研究所的ABI Prism 7000 SDS software 1.5进行相对定量分析,PCR反应结束后自动生成报告,采用文献中相对定量 2-△△Ct法[15]计算目的基因的相对表达量。用SPSS 16.0对数据进行统计分析,并用LSD法和Duncan氏法进行多重比较,结果用平均值±标准差表示。
2 结果
2.1 背最长肌剪切力及 CAST、CaN、CaM 和NFAT m RNA表达量
由表3可知,12%组肌肉剪切力显著低于20%组(P<0.05),16%组与其他组差异不显著(P>0.05),肌肉剪切力随饲粮蛋白质水平升高而增加,即蛋白水平越高肌肉嫩度越差。20%组的CAST和CaN mRNA表达量极显著高于12%组和16%组(P<0.01),12%组与16%组差异不显著(P>0.05),CAST和 CaN mRNA在背最长肌中的表达量随饲粮蛋白水平升高而增加;NFAT和CaM mRNA表达量在3个处理组间差异不显著(P >0.05)。

表3 饲粮蛋白质水平对猪背最长肌剪切力、CaN、NFAT、CaM和CAST m RNA相对表达量的影响Table 3 Effects of dietary protein level on the shear force and mRNA relative expression levels of CaN,NFAT ,CaM and CAST of longissimus dorsi in finishing pigs
同行数据肩标不同小写字母表示差异显著(P<0.05),不同大写字母表示差异极显著(P<0.01),相同字母表示差异不显著(P >0.05)。
In the same row,valuesw ith different small superscriptsmean significant difference(P <0.05),and w ith different capital letter superscriptsmean significant difference(P <0.01),while w ith the same letter superscriptsmean no significant difference(P >0.05).
2.2 背最长肌剪切力与 CaN、NFAT、CaM 和CAST m RNA表达量相关性
由表4可知,背最长肌剪切力与CAST mRNA表达量极显著正相关(P<0.01),相关系数为0.713;背最长肌剪切力与 CaN、CAM 和 NFAT mRNA表达量的相关系数分别为0.439、0.135和0.374,无显著相关(P > 0.05)。CaN、CaM 与CAST mRNA表达量显著正相关(P<0.05),相关系数分别为 0.594、0.596;NFAT 与 CAST mRNA表达量的相关系数为0.376,无显著相关(P>0.05)。NFAT、CaM 与CaN mRNA表达量显著正相关(P <0.05),相关系数分别为 0.536、0.546。CaM与NFAT mRNA表达量显著正相关(P<0.05),相关系数为 0.623。
3 讨论
3.1 饲粮蛋白质水平对背最长肌剪切力的影响
嫩度是肉品质的一种感官特征,也是其食用质量与商业价值的重要指标。它来源于肌束的结构和生化特性,特别是肌原纤维和中等细丝的结构和生化特性,以及肌间结缔组织。肉的嫩化是尸僵发生后一定时间内,肌纤维碎裂、肌肉重新变软、嫩度增加的过程[16]。肌肉嫩度的评定通常采用肉的咬力值和剪切力来表示。Goerl等[17]研究了饲粮蛋白质水平对28~104 kg猪肉品质的影响,同样发现随饲粮蛋白质水平的增加胴体背膘厚下降、瘦肉率增加、肉嫩度下降。Davey等[18]研究发现,用高蛋白质饲粮饲喂瘦肉型猪,可提高瘦肉率、降低肌肉内脂肪水平、降低肉的嫩度。张克英等[19]报道生长肥育猪60 kg后饲粮采用理想蛋白质水平12%,可获得较好的猪肉品质,在此基础上,适当降低饲粮蛋白质水平有利于改善猪肉的风味和嫩度,但进一步提高蛋白质水平,则可能影响猪肉品质,这与本试验的结果相似,随着饲粮蛋白质水平的升高,剪切力值上升,表明提高蛋白质水平对改善肌肉的嫩度有着负面影响。
3.2 饲粮蛋白质水平对背最长肌CAST和CaNNFAT信号途径相关蛋白m RNA表达量的影响
钙蛋白酶系统由一系列的钙蛋白酶和CAST构成。钙蛋白酶是肌原纤维蛋白质降解过程中的关键酶。CAST可抑制钙蛋白酶的活性,当钙蛋白酶被Ca2+激活后,如果附近有CAST存在,将迅速与之结合,影响其自溶稳定性,抑制钙蛋白酶的活性,从而保证钙蛋白酶只进行局部的特定位点的水解。Thomson等[20]报道,营养水平对断奶阉公羊骨骼肌钙蛋白系统活性均无影响,但肌肉蛋白质沉积速度与CAST活性呈正相关,肌肉蛋白质降解速度与钙激活酶Ⅰ(μ-calpain)活性呈负相关。用不同蛋白质水平的饲粮饲喂56只狗,经过70 d后检测得到CAST在狗的骨骼肌中表达量随着蛋白质水平的提高而呈上升趋势[21]。本试验结果表明,饲粮蛋白质水平与背最长肌CAST mRNA表达量有一定的相关,随着蛋白质水平的升高背最长肌CAST mRNA表达量增加,反映了蛋白质水平的提高可加快肥育猪的蛋白质沉积,从而抑制钙蛋白酶对肌原纤维蛋白质的降解。这一结论与 Thomson 等[20]在羊的试验报道和 Helman 等[21]在狗的试验报道一致。

表4 猪背最长肌剪切力、CaN、NFAT、CaM和CAST m RNA相对表达量回归方程Table 4 Regression equations of shear force and mRNA relative expression levels of CaN,NFAT,CaM and CAST of longissimus dorsi in finishing pigs
CaN-NFAT信号途径是调节骨骼肌生长,决定肌纤维类型特征的主要信号途径。本课题组曾研究了不同饲喂方式以及不同饲粮蛋白质水平对猪骨骼肌中CAST和钙蛋白酶mRNA表达量的影响[13-14],而就饲粮蛋白质水平对 CaN-NFAT 信号途径相关蛋白mRNA表达量的影响国内外鲜有报道。本试验中,随饲粮蛋白质水平的升高猪背最长肌 CaN mRNA表达量显著增加,NFAT、CaM mRNA表达量随CaN mRNA表达量增加而增加,且三者之间的变化相关性明显,其原因可能是当动物摄入蛋白质量增加,通过神经刺激使得胞质内 Ca2+浓度升高、CaN被激活,可以让下游的NFAT家族成员脱磷酸并转入细胞核,与细胞内转录因子结合,使得CaM表达增加。由于CAST在不同类型纤维中的表达是不同的[22],CAST的表达与CaN-NFAT信号途径的关系如何是一个值得研究的课题。但目前这方面的研究报道极少,本试验的结果可为进一步研究CAST与CaN-NFAT信号途径之间的调控机理提供参考。
3.3 背最长肌CAST和CaN-NFAT信号途径对嫩度的影响
影响肉的嫩度的主要因素有肌节长度、结缔组织含量及肌肉结构蛋白水解敏感性[23]。肉的嫩度是肌肉内部结构的反映,在一定程度上反映了肌原纤维、结缔组织以及肌肉脂肪的含水量、分布和化学结构[24]。CAST对钙蛋白酶具有抑制作用,其分解产物对钙蛋白酶也具有抑制作用,而钙蛋白酶对肌肉纤维具分解作用,因此理论上它可以作为嫩度的候选基因。Koohmaraie等[25]研究认为,高水平的CAST抑制宰后肌肉蛋白质水解和嫩化。孙立彬等[26]对125头猪进行屠宰,通过CAST基因型与屠体性状、肉质性状的相关性分析,得出CAST基因对肌内脂肪含量具有显著影响。武艳群等[27]试验进一步证明CAST是与肉质相关性非常高的候选基因。本试验结果同样证实CAST与肌肉剪切力存在显著相关性,CAST mRNA表达量的增加降低了猪背最长肌的嫩度。
随着肉质研究的发展,人们开始关注肌纤维类型的特性及其分布规律,大量研究表明,肌纤维类型是决定肉品质的重要因素[28-30]。CaN-NFAT信号途径是调节不同纤维类型基因表达的关键机制,小鼠CaN基因敲除模型证实CaN-NFAT信号途径可激活骨骼肌慢缩型纤维的基因表达[31]。对CaN-NFAT信号途径而言,Ca2+浓度持续增高比短暂脉冲式增高更重要[32],这意味着在快缩纤维和慢缩纤维中,不同的Ca2+环境所诱发的CaNNFAT信号系统反应程度不同。CaN-NFAT信号通路被激活后,可以减轻肌营养不良鼠(mdx鼠)肌肉萎缩病变的程度[33]。CaN可以促进骨骼肌组织的再生。Sakuma等[34]研究发现,在大鼠的再生肌肉组织中,CaN活性及去磷酸化的NFAT含量均大幅度增加,得出Ca2+-CaN-NFAT信号途径在肌肉的发生、发育及功能维持中非常重要。本试验中,CaN、NFAT、CaM mRNA表达量均随剪切力的升高而呈增加趋势,说明CaN-NFAT信号途径可能通过调节肌纤维的类型而在肌肉剪切力增高中起一定作用。对于CaN-NFAT信号途径与肌肉嫩度的研究国内外鲜见报道,能否通过CaN-NFAT信号途径相关蛋白的表达来进一步调控肉品质还有待进一步的研究。
4 结论
①饲粮高蛋白质水平显著降低背最长肌嫩度、提高CAST和CaN mRNA表达量。
②背最长肌嫩度与CASTmRNA表达量正相关,CaN-NFAT信号途径无相关性。
[1] KAO S C,WU H,XIE J M,et al.Calcineurin/NFAT signaling is required for neuregulin-regulated schwann cell differentiation[J].Science,2009,323(5914):651-654.
[2] KOOHMARAIEM.Biochem ical factors regulation the toughening and tenderization process of meat[J].Meat Science,1996,43:S193.
[3] MARTINS P A,SALIC K,GLADKA M M,et al.M icroRNA-199b targets the nuclear kinase Dyrk1a in an auto-amplification loop promoting calcineurin/NFAT signaling[J].Nature Cell Biology,2010,12:1220-1227.
[4] ABDUL H M,SAMA M A,FURMAN J L,et al.Cognitive decline in A lzheimer’s disease is associated w ith selective changes in calcineurin/NFAT signaling[J].The Journal of Neuroscience,2009,29(41):12957-12969.
[5] COURTWRIGHT A,REIHANIS S,ARBISER JL,et al.Secreted frizzle-related protein 2 stimulates angiogeneses via a calcineurin/NFAT signaling pathway[J].Current Issue,2011,71(1):471 -484.
[6] DONNELL S M,THOMAS W,KUBO H,et al.CaMKⅡnegatively regulates calcineurin/NFAT signaling in cardiac m yocytes[J].Circulation Research,2009,105(4):316-325.
[7] HOUSER S R,MOLKENTIN JD.Does contractile Ca2+control calcineurin/NFAT signaling and pathological hypertrophy in cardiac myocytes[J].Science Signaling,2008,25(1):1126 -1130.
[8] PALMER B R,ROBERTS N,HICKFORD JG H,et al.Rapid communication:PCR-RFLP for MspⅠand NcoⅠ in the ovine calpastatin gene[J].Journal of Animal Science,1998,76:1499-1450.
[9] CIOBANU D C,BASTIAANSEN JW M,LONERGAN SM,et al.New alleles in calpastatin gene are associated w ith meatquality traits in pigs[J].Journal of Animal Science,2004,82:2829-2838.
[10] KURYL J,KAPELANSKIW,PIERZCHALA M.Prelim inary observations on the effect of calpastatin gene(CAST)polymorphism on carcass traits in pigs[J].Animal Science Papers and Reports,2003,21:87-95.
[11] COCKETT N E,SHAY T L,GREEN R D,et al.Rapid communication:a TaqⅠrestriction fragment length polymorphism in the bovine calpastatin gene[J].Journal of Animal Science,1995,73(12):3790.
[12] BUCHHOLZ M,SCHATZ A,WAGNER M,et al.Overexpression of c-myc in pancreatic cancer caused by ectopic activation of NFATc1 and the Ca2+/calcineurin signaling pathway[J].European Molecular Biology Organization,2006,25:3714-3724.
[13] 张勇,高彦,朱宇旌,等.不同饲喂方式对猪背最长肌钙蛋白酶抑制蛋白和钙蛋白酶基因表达及剪切力的影响[J].动物营养学报,2010,22(3):640-646.
[14] 张勇,李方方,朱宇旌,等.日粮不同蛋白质水平对猪骨骼肌钙蛋白酶抑制蛋白和钙蛋白酶基因表达及嫩度的影响[J].动物营养学报,2008,20(3):360-365.
[15] LIVAK K J,SCHM ITTGEN T D.Analysis of relative gene expression data using real-time quantitative PCR and the 2-△△Ctmethod[J].Method,2001,25(4):402-408.
[16] 翟峰,张勇.钙蛋白酶抑制蛋白与肌肉嫩度相关性[J].中国饲料,2007,16:11 -13.
[17] GOERL K F,EILERT S J,MANDIGO RW,et al.Pork characteristics as affected by two populations of sw ine and six crude protein levels[J].Journalof Animal Science,1995,73:3621-3626.
[18] DAVEY R J.Grow th and carcass characteristics of high-and low-fat sw ine fed diets varying in protein and lysine content[J].Journal of Animal Science,1976,43:598-605.
[19] 张克英,陈代文,罗献梅.饲粮理想蛋白水平对猪肉品质的影响[J].四川农业大学学报,2002(1):37-39.
[20] THOMSON B C,HOSKING B J,SAINZ R D,et al.The effect of nutrional status on protein degradation and components of the calpain system in skeletal muscle of weaned wether lambs[J].The Journal of Agricultural Science,1997,129:471-477.
[21] HEMAN E E, HUFF-LONERGAN E, DAVENPORT G M,et al.Effcet of dietary protein on calpastatin in CaNine skeletalmuscle[J].Journal of Animal Science,2003,81:2199-2205.
[22] PARR T,JEWELL K K,SENSKY P L,et al.Expression of calpastatin isoforms in muscle and functionality of multiple calpastatin promoters[J].Archives of Biochem istry and Biophysics,2004,427:8-15.
[23] HIAN M A,MORTON JD,KENT M P.Intermuscular variation in tenderness:association w ith the ubiquitous and muscle-specific calpains[J].Journal of Animal Science,2001,79(1):122-l32.
[24] 李德发.营养调控肉品质量的研究现状及发展趋势[M]//动物营养研究进展.北京:农业科技出版社,2004:7-14.
[25] KOOHMARAIE M,GEESINK G H.Contribution of postmortem muscle biochem istry to the delivery of consistent meat quality w ith particular focus on the calpain system[J].Meat Science,2006(74):34 -43.
[26] 孙立彬,孟和,王起山,等.猪2型钙蛋白酶抑制蛋白基因cDNA的克隆序列分析[J].上海交通大学学报:农业科学版,2005,23(3):66 -69.
[27] 武艳群,吴旧生,赵晓枫,等.猪 CAST基因与肌纤维组织学特性及屠宰性状的相关性分析[J].遗传,2007,29(1):65 -69.
[28] KLONT R E,EIKELNBOOM L B.Muscle fibre type and meat quality[J].Meat Science,1998,49(Suppl.1):S219 - S229.
[29] KARLSSON A H ,KLONT R E,FERNANDEZ X.Skeletalmuscle fibres as factors for pork quality[J].Livestock Production Science,1999,60:255-269.
[30] CHANG K C,DA COST N,BLACKLEY R,et al.Relationships of myosin heavy chain fibre types to meat quality traits in traditional and modern pigs[J].Meat Science,2003,64:93-103.
[31] PARSONS S A,M ILLAY D P,W ILKINS B J,et al.Genetic loss of calcineurin blocksmechanicaloverload-induced skeletal muscle fiber type sw itching but not hypertrophy[J].Journal of Biological Chemistry,2004,279(25):26192-26200.
[32] TIMMERMAN L A,CLIPSTONE N A,HO SN,et al.Rapid shuttling of NFAT in discrim ination of Ca2+signals and immunosuppression[J].Nature,1996,383(6603):837-840.
[33] CHAKKALAKAL J V,HARRISON M A,CARBONETTO S,et al.Stimulation of calcineurin signaling attenuates the dystrophic pathology in mdx m ice[J].Human Molecular Genetics,2004,13(4):379-388.
[34] SAKUMA K,NISHIKAWA J,NAKAO R,et al.Calcineurin is a potent regulator for skeletalmuscle regeneration by association w ith NFATc1 and GATA-2[J].Acta Neuropathologica,2003,105(3):271 -280.
猜你喜欢
橡塑技术与装备(2022年10期)2022-10-03
动物营养学报(2022年8期)2022-09-01
水利科技与经济(2021年11期)2021-12-04
中国饲料(2021年17期)2021-11-02
肉类研究(2017年10期)2017-11-20
中国农业科学(2016年17期)2016-12-01
浙江大学学报(工学版)(2016年2期)2016-06-05
浙江农业科学(2016年11期)2016-05-04
动物营养学报(2015年9期)2016-01-07
动物营养学报(2012年4期)2012-09-20
