
无性型虫草菌株qsun-1的生长特性
2011-09-27 01:51张晓君,孙庆元,王建军
大连工业大学学报 2011年5期
张 晓 君, 孙 庆 元, 王 建 军
( 大连工业大学 生物工程学院, 辽宁 大连 116034 )
0 引 言
虫草菌[Cordycepssinensis(Berkey) Sacc.]隶属于真菌界(Fungi)、子囊菌门(Ascomycota)、子囊菌纲(Ascomycetes)、粪壳菌亚纲(Sordariomycetidae)、肉座菌目(Hypocreales)、麦角菌科(Clavicipitaceae)、虫草属(Cordyceps)[1-3]。一般虫草菌侵染寄主后,菌丝体分化形成垫状结构即子座。子座膨大并产生子实体,即子囊壳,成熟的子囊壳中产生子囊和子囊孢子。在野外虫草菌孢子在夏季萌发侵染土壤中的蝙蝠蛾幼虫后,形成的子座在次年5月份长出地面[4],貌似草,这种蝙蝠蛾幼虫与虫草菌的复合体称为冬虫夏草。冬虫夏草主要分布于我国的青藏高原海拔3 500~5 000 m的高山草甸或高山灌丛草甸,在四川、云南和甘肃也有少量分布[5]。作者从野外采集的冬虫夏草中分离无性型虫草菌株,对菌虫体组织和子座组织萌发、无性型虫草菌株菌落形态及显微结构进行了观察,同时对无性型虫草菌株发酵培养基组成成分进行了优化,以期为人工培育冬虫夏草以及无性型虫草菌株发酵提供理论基础。
1 实验材料
1.1 冬虫夏草
新鲜冬虫夏草,采自四川省阿坝藏族羌族自治州黑水县,冰箱中4 ℃保存。
1.2 培养基
分离培养基(PDA培养基):马铃薯20%,葡萄糖2%,VB10.01%,KH2PO40.1%,MgSO40.1%,琼脂2%,pH 6.0。用于无性型虫草菌株的分离。
斜面培养基(1%牛肉粉PDA培养基):马铃薯20%,葡萄糖2%,牛肉粉1%,VB10.01%,KH2PO40.1%,MgSO40.1%,琼脂2%,pH 6.0。用于无性型虫草菌株的保藏。
基础培养基:葡萄糖3%,牛肉粉2%,KH2PO40.1%,MgSO40.1%,VB10.01%,pH 6.0。用于无性型虫草菌株的发酵。
2 实验方法
2.1 分 离
用水清洗冬虫夏草表面的杂质,在无菌条件下分别用0.1%氯化汞、75%酒精对冬虫夏草进行表面消毒,无菌水冲洗多次[6],以头部为界将其剪开,生长子座的头部称为子座组织,头部以外的称为菌虫体组织。将子座及菌虫体组织分别放入分离培养基中,在18 ℃培养,观察,重复3次,统计萌发情况,并且挑取萌发的菌丝体进一步分离纯化后接入斜面培养基保藏。
2.2 生长观察
将分离的无性型虫草菌株qsun-1分别接入PDA及1%牛肉粉PDA平板中,18 ℃培养,观察生长情况。同时将接有无性型虫草菌株qsun-1的PDA培养基放在15、18、25、32 ℃培养,观察生长情况。以菌落直径作为衡量生长情况的指标,每隔5 d测量1次。
2.3 显微结构观察
无菌操作,用接种棒从液体培养基中挑取菌丝,放在滴有纯化水的载玻片上,轻轻将菌丝展开,不染色直接用显微镜进行观察[7]。
2.4 发酵培养基的优化
将无性型虫草菌株qsun-1在1%牛肉粉PDA液体培养基中活化3 d,以5%的接种量接入发酵培养基。发酵条件:装液量每250 mL装100 mL,温度18 ℃,摇床转速120 r/min,时间5 d。发酵结束后3 000 r/min离心10 min分离菌丝体,烘干称重。
以葡萄糖、果糖、蔗糖、可溶性淀粉及甘露醇为碳源,确定最佳碳源。确定蔗糖为最佳碳源后,分别考察1%、2%、3%、4%、5%蔗糖对无性型虫草菌株qsun-1菌体生长的影响,确定最佳蔗糖质量分数。以牛肉粉、蛋白胨、NaNO3、(NH4)2SO4及尿素为氮源,确定最佳氮源。确定蛋白胨为最佳氮源后,分别考察0.5%、1.0%、1.5%、2.0%、2.5%蛋白胨对无性型虫草菌株qsun-1菌体生长的影响,确定最佳蛋白胨质量分数。上述条件确定后依次考察0.05%、0.10%、0.15%、0.20%、0.25% MgSO4和KH2PO4对无性型虫草菌株qsun-1菌体生长的影响,确定最佳的MgSO4、KH2PO4的质量分数[8-10]。
3 结果与分析
3.1 萌发和分离
对新鲜冬虫夏草不同部位进行分离,得到无性型虫草菌株qsun-1(表1)。由表1可以看出,冬虫夏草子座组织的菌体萌发率为60%,而菌虫体组织的菌体萌发率仅达到14%,可见子座组织菌丝体容易萌发,而菌虫体组织可能由于易受杂菌污染而使萌发受阻。因此,作者从冬虫夏草子座中培养并分离了无性型虫草菌株。

3.2 生长特点
3.2.1 菌落形态
无性型虫草菌株qsun-1在PDA和1%牛肉粉PDA平板中生长10 d后菌落形态有所不同(图1)。在PDA培养基中菌落稍微隆起,中心凹陷并有少量水珠,灰白色,菌丝较短,菌落周围有一透明圈[图1(a)],菌落背面暗黄色[图1(b)],菌落直径21 mm。在1%牛肉粉PDA培养基中菌落明显隆起,纯白色,菌丝粗壮[图1(c)],菌落背面淡黄色[图1(d)],菌落直径30 mm。

图1 菌株qsun-1菌落形态
3.2.2 菌落生长情况
无性型虫草菌株属于嗜低温菌株,对温度变化反应显著。在不同温度下培养的无性型菌株qsun-1生长状况也不同(表2)。无性型虫草菌株qsun-1在15~18 ℃生长良好,在25 ℃生长受到抑制, 在32 ℃条件下未发现菌株生长。
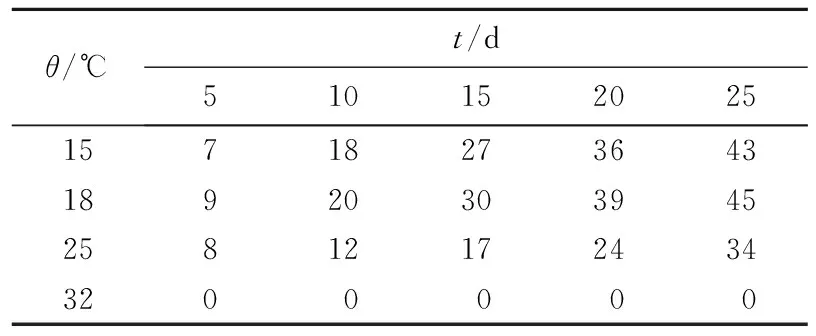
表2 不同温度下菌株qsun-1菌落直径
3.3 显微结构
对无性型虫草菌株qsun-1液体培养物进行显微观察,从显微结构可以看到液体培养的菌丝主要为管状菌丝,菌丝有较明显的分隔,粗细均匀,每节菌丝均透明(图2)。
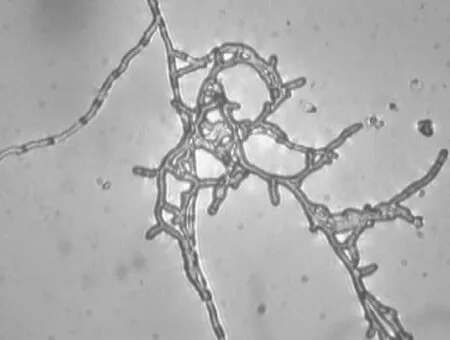
图2 菌株qsun-1显微结构(×250)
3.4 发酵培养基的优化
3.4.1 碳源对qsun-1生长的影响
无性型虫草菌株qsun-1可以利用多种碳源。当蔗糖为碳源时生长最好,菌体产量达到8.0 g/L,其次是果糖、甘露醇、淀粉和葡萄糖(表3)。

3.4.2 蔗糖质量分数对qsun-1生长的影响
无性型虫草菌株qsun-1在蔗糖为1%时生长较慢;在蔗糖质量分数达到2%时,生长量最大; 当蔗糖质量分数超过2%时,菌体生长受到一定程度的抑制;当蔗糖质量分数达到5%时,菌体生长明显受阻(图3)。
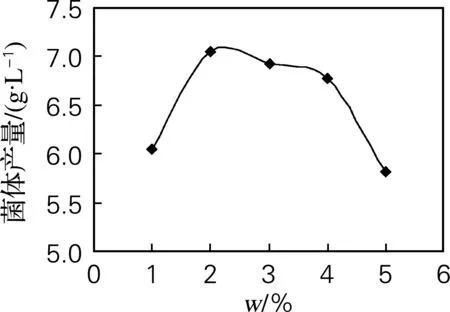
图3 蔗糖质量分数对菌株qsun-1生长的影响
3.4.3 氮源对qsun-1生长的影响
无性型虫草菌株qsun-1可利用多种氮源。当以蛋白胨为氮源时生长最好,菌体产量达到8.1 g/L,其次为NaNO3、牛肉粉、(NH4)2SO4。以尿素为氮源时,未见无性型虫草菌株qsun-1生长(表4)。

3.4.4 蛋白胨质量分数对qsun-1生长的影响
无性型虫草菌株qsun-1在蛋白胨为0.5%时,生长缓慢;随着蛋白胨质量分数的增加生长量逐渐增加,在蛋白胨质量分数达到2%时,菌体生长达到最大值;蛋白胨质量分数超过2%后,菌体生长受到抑制(图4)。
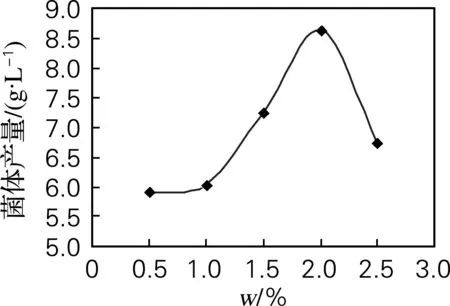
图4 蛋白胨质量分数对菌株qsun-1生长的影响
3.4.5 MgSO4质量分数对qsun-1生长的影响
无性型虫草菌株qsun-1在MgSO4为0.05%时生长较慢,在MgSO4达到0.1%时生长量达最大;MgSO4超过0.1%后生长受到一定程度的抑制,并随着MgSO4浓度的增加而抑制程度增加(图5)。

图5 MgSO4质量分数对菌株qsun-1生长的影响
3.4.6 KH2PO4质量分数对qsun-1生长的影响
无性型虫草菌株qsun-1在KH2PO4为0.05%时生长较慢,随着KH2PO4浓度的增加生长量逐渐增加,在0.2%左右时达到最大值,在超过0.2%后菌体生长受到抑制(图6)。

图6 KH2PO4质量分数对菌株qsun-1生长的影响
4 结 论
冬虫夏草子座组织菌丝体容易萌发,而菌虫体组织中可能由于易受杂菌污染菌丝体萌发受阻。作者从冬虫夏草子座中培养并分离出无性型虫草菌株qsun-1。通过在PDA和1%牛肉粉PDA中培养无性型虫草菌株qsun-1,发现1%牛肉粉PDA更适于qsun-1生长,qsun-1在1%牛肉粉PDA培养10 d,菌落明显隆起,纯白色,菌丝粗壮,菌落背面淡黄色,菌落直径30 mm。观察qsun-1的显微结构,可以看到液体培养的菌丝主要为管状菌丝,菌丝有较明显的分隔,粗细均匀,每节菌丝均透明。通过在不同温度下培养菌株qsun-1,发现其生长温度范围在15~18 ℃。无性型虫草菌株qsun-1最佳发酵培养基组成为:蔗糖2%,蛋白胨2%,MgSO40.1%,KH2PO40.2%。
[1] 薛召东,杨瑞林,易永健. 冬虫夏草及其研究与利用[J]. 中国麻业, 2002, 24(3):50-51.
[2] 李进,冯成强,张文生. 冬虫夏草研究回顾与展望[J]. 中国农学通报, 2008, 24(2):382-383.
[3] 魏镜超. 真菌鉴定手册[M]. 上海:上海科学技术出版社, 1979:218-219.
[4] 吕献康,沈建华,舒小英. 冬虫夏草生态生物学特性考察报告[J]. 中国现代应用药学杂志, 2005, 22(2):134-135.
[5] 王丽娟. 冬虫夏草资源的分布调查[J]. 甘肃林业, 2007(4):40-41.
[6] 王忠,马启龙,乔正强. 甘肃冬虫夏草菌分离培养研究[J]. 甘肃农业科技, 2001(7):43-44.
[7] 刘彦威,苏敬良,韩博,等. 冬虫夏草无性型的分离培养和菌丝显微结构的观察[J]. 黑龙江畜牧兽医, 2005(10):67-69.
[8] 赵润,郭成金. 冬虫夏草菌丝体液体培养基的优化[J]. 天津师范大学学报:自然科学版, 2008, 28(1):8-11.
[9] XIAO Jian-hui, CHEN Dai-xiong, LIU Jin-wei, et al. Optimization of submerged culture requirements for the production of mycelial growth and exopolysaccharide byCordycepsjiangxiensisJXPJ 0109[J]. Journal of Applied Microbiology, 2004, 96(5):1105-1116.
[10] KIM S W, HWANG H J, XU C P, et al. Optimization of submerged culture process for the production of mycelial biomass and exo-polysaccharides byCordycepsmilitarisC738[J]. Journal of Applied Microbiology, 2003, 94(1):120-126.
猜你喜欢
中老年保健(2022年2期)2022-08-24
当代水产(2022年1期)2022-04-26
中老年保健(2021年12期)2021-11-30
发明与创新·小学生(2021年12期)2021-01-06
军事文摘(2020年22期)2021-01-04
阿来研究(2018年1期)2018-07-10
中国调味品(2017年2期)2017-03-20
汉语世界(2016年3期)2016-11-16
现代检验医学杂志(2016年3期)2016-11-15
中央民族大学学报(自然科学版)(2016年3期)2016-06-27
