
定量研究菲啶对酵母朊病毒[PSI+]的治愈作用
2012-02-10 01:20钟正伟王莉洁谢辉李辉何剑为宋有涛
生物工程学报 2012年6期
钟正伟,王莉洁,谢辉,李辉,3,何剑为,3,宋有涛,3
1 辽宁大学生命科学院,辽宁 沈阳 110036
2 承德石油高等专科学校化学工程系,河北 承德 067000
3 辽宁省动物资源与疫病防治重点实验室,辽宁 沈阳 110036
朊病毒是一类能在哺乳动物中引起疯牛病、绵羊瘙痒症和人克雅氏病等可传染性海绵样脑病的病原体,迄今为止对于朊病毒疾病仍缺乏有效的治疗药物[1-2]。近年来,利用抗朊病毒药物筛选模型筛选抗朊病毒候选药物是国际上的研究热点,研究者通过构建动物模型、动物细胞模型、无细胞模型和酵母模型在抗朊病毒候选药物的筛选方面进行了大量研究,获得了戊聚糖多硫酸酯、刚果红、四磺酸基酞菁、两性霉素B和奎纳克林等一系列具有抗朊病毒作用效果的候选药物[3-9]。
由于酵母朊病毒[PSI+]具有易于检测的遗传表型 (红色为正常型,白色为致病型) 及其与动物具有严格种间屏障的优势,因此基于酵母细胞的抗朊病毒候选药物筛选模型在大规模筛选上具有经济、安全和易实现高通量筛选等特性而越来越受到关注[5-10]。利用酵母朊病毒的遗传表型并结合蛋白水平的 SDS-PAGE/Western blotting分析,Bach等从近4 000种小分子化合物中筛选发现了菲啶 (Phenanthridine,Phen)、6-氨基菲啶(6-Aminophenanthridine) 等 11种化合物可以使酵母朊病毒的致病型白色表型转换为正常型红色表型,同时其淀粉样聚集状态可以转换为可溶单体状态[5]。值得注意的是,菲啶衍生物6-氨基菲啶作用酵母朊病毒[PSI+]后产生了93%的介于红色的阳性表型与白色的阴性表型之间的粉色中间表型,该表型的朊病毒仍为聚集状态,并在传代实验中遗传不稳定,多次传代后转变为白色表型[5]。而在此前的研究中,盐酸胍等化合物虽然能够治愈[PSI+]却不能引起酵母细胞粉色表型的出现[11]。
近年来的研究表明,朊病毒聚集体的大小较聚集状态更能够反映朊病毒“种子”(Seeds) 的聚集和传播能力[12-14]。这暗示着在蛋白和细胞水平对朊病毒聚集体大小的定量分析可以更准确地评价抗朊病毒候选药物的作用效果,尤其是对粉色中间表型而言。因此,本研究利用本实验室构建的新型抗朊病毒药物筛选酵母模型,即基因重组的荧光标签 GFP插入 Sup35 (NGMC) 的[GPSI+]酵母菌株,引入荧光漂白后恢复技术(Fluorescence redistribution after photobleaching,FRAP) 和半变性琼脂糖凝胶电泳 (Semidenaturing detergent-agarose gel electrophoresis,SDD-AGE) 技术分别在细胞水平和蛋白水平定量分析了菲啶作用后酵母细胞内朊病毒聚集体的大小变化[15-17]。另外,本研究还系统研究了菲啶作用后产生的粉色表型酵母中朊病毒的聚集状态。这些实验结果弥补了Bach等对菲啶等化合物的抗酵母朊病毒效果缺乏细胞和蛋白水平定量分析的缺陷。更重要的是,该研究成果为基于酵母模型的抗朊病毒药物次级筛选也提供了一套精确的定量方法。
1 材料与方法
1.1 材料
1.1.1 菌株
野生型酵母 779-6A (NMC):MATα kar1 SUQ5 ade2-1 his3Δ202 leu2Δ1 trp1Δ63 ura3-52 sup35:: KanMX/ pJ501,[PSI+],由美国国立卫生院 (NIH) Masison DC. 博士惠赠。重组型酵母(NGMC):MATα kar1 SUQ5 ade2-1 his3Δ202 leu2Δ1 trp1Δ63 ura3-52 sup35 :: KanMX/pJ510,[GPSI+],GFP基因插入SUP35基因的第123~124位之间,由本实验室构建。将[PSI+]或[GPSI+]细胞划线至含有5 mmol/L盐酸胍的1/2YPD固体平板上,培养7 d后,分离红色单菌落得到[psi-]或[Gpsi-]细胞。
1.1.2 培养基
1/2YPD培养基:酵母浸粉0.5%,蛋白胨2%,葡萄糖2%,固体平板加入2%琼脂。
YPAD培养基:酵母浸粉1%,蛋白胨2%,葡萄糖2%,腺嘌呤0.04%,固体平板加入2%琼脂。
YPD培养基:酵母浸粉1%,蛋白胨2%,葡萄糖2%,固体平板加入2%琼脂。
1.1.3 主要试剂和仪器
盐酸胍、菲啶、刀豆球蛋白购于Sigma公司,兔抗GFP抗体与HRP标记的羊抗兔IgG购于金斯特生物科技有限公司,其他试剂均为国产分析纯。琼脂糖水平电泳槽 (DYCO-31DN,北京六一公司)、转膜仪 (Bio-Rad公司,美国)、珠磨破碎仪 (Biospec公司,美国)、激光共聚焦显微镜(OLYMPUS FV1000S-SIM/IX81,中国医科大学实验中心)。
1.2 方法
1.2.1 遗传表型实验
[PSI+]和[GPSI+]酵母细胞在 30 ℃过夜活化培养后,调整菌液OD600至0.2左右,按10%接种量分别接种到含有指定化合物的1/2YPD液体培养体系,所有体系于24 ℃振荡培养5 d,在培养时间内间隔24 h更换培养基和化合物[5]。每天取培养液稀释后涂布于1/2YPD固体平板,平板于24 ℃培养7 d,观察菌落颜色变化,统计不同颜色菌落的比例。
1.2.2 荧光焦点 (Foci) 和FRAP实验
将化合物作用[GPSI+]酵母细胞1~5 d后涂布培养的1/2YPD固体平板上出现的不同颜色菌落接种到YPAD液体培养基中30 ℃培养2 d后,取少量菌液滴在干燥的覆盖有2 g/L刀豆球蛋白溶液的洁净载玻片上,加盖玻片并封片,在100倍共聚焦荧光显微镜 (OLYMPUS-FV1000型)下观察朊病毒在酵母活细胞内形成荧光焦点的情况。随后利用荧光漂白后恢复技术在活细胞中分析酵母朊病毒聚集体的迁移速率。FRAP技术的原理是采用强激光在短时间内将荧光分子漂白区域的荧光淬灭,由于活细胞内分子处于流动状态,因此随着漂白区域外的荧光分子逐渐流入,漂白区内的荧光会逐渐恢复。通过测定漂白区域荧光恢复的速率可以获得荧光分子迁移速率的相关信息[16,18]。FRAP实验参考文献的方法并作适当修正[16],激发波长为 488 nm,输出功率为最大输出功率的2%,针孔调整为3 μm,物镜为40 倍,扫描整个样品,选取漂白区 (ROI),漂白功率为100%,漂白程度为50%~80%,漂白后采集数据时间间隔设定为0.5 s,采集次数为40~50 次,漂白结束后,保存数据。按照公式FR(t)=(Ft−Fmin)/(Fi−Fmin) 计算相对荧光强度,其中Ft表示细胞漂白区在t时间的荧光强度,Fi表示t=0时间的荧光强度,Fmin表示细胞漂白后的最小荧光强度,FR(t)表示t时间细胞的相对荧光强度。以FR(t)VS t作图得到荧光漂白后恢复曲线。
1.2.3 SDD-AGE/Western blotting实验
将化合物作用[GPSI+]酵母细胞1~5 d后涂布培养的1/2YPD固体平板上出现的不同颜色菌落活化后接种到 YPD液体培养基中 30 ℃培养约12 h,到菌液 OD600=1.5左右时停止培养,离心收集细胞,然后加入裂解缓冲液,用珠磨仪裂解细胞,离心收集上清得到蛋白样品;SDD-AGE/ Western blotting实验流程参考文献的方法并作适当调整[17],进行1.5%的琼脂糖凝胶电泳,电泳结束后在0 ℃、100 V条件下转膜1 h,将蛋白转移到PVDF膜上,兔抗-GFP抗原 (1∶2 000) 孵育1 h,4 ℃过夜,HRP标记的羊抗兔IgG (1∶5 000)孵育1 h,采用ECL发光试剂盒对其进行放射自显影检测,获得图像信息。
2 结果与分析
2.1 菲啶对酵母朊病毒[PSI+]表型的影响
酵母朊病毒[PSI+]是翻译终止因子 Sup35p的淀粉样聚集形式[19]。在[PSI+]细胞中,Sup35p主要以聚集体形式存在导致无义突变抑制产生终止密码子通读现象,使ade2-1突变的[PSI+]细胞在腺嘌呤缺乏的培养基上生长为白色菌落;在被治愈的[psi-]细胞中,其Sup35p主要以可溶形式存在,非无义抑制的ade2-1突变株需要在含有腺嘌呤的培养基上生长,细胞由于累积腺嘌呤合成途径中的红色中间产物而形成红色菌落,这个颜色指示系统可用来初步分析酵母朊病毒的存在状态[13]。鉴于在Bach的研究中,菲啶治愈野生型酵母朊病毒必须存在增效剂亚治愈浓度的盐酸胍 (0.5 mmol/L,在此浓度下盐酸胍可以通过部分竞争性抑制 Hsp104的活性削弱[PSI+]的强度而不改变[PSI+]的性质)[5],为此我们在0.5 mmol/L盐酸胍条件下研究菲啶对酵母朊病毒的治愈作用。如表1所示,阴性对照0.5 mmol/L盐酸胍和阳性对照5 mmol/L盐酸胍对酵母朊病毒[GPSI+]的5 d治愈率分别为0%和94.8%,而0.2 mmol/L菲啶在增效剂存在情况下治愈率为84.6%,与相同条件下菲啶和盐酸胍对野生型[PSI+]的治愈率基本相似并略高。这暗示着在基于酵母的抗朊病毒药物筛选过程中,可以用酵母朊病毒NGMC菌株代替野生型酵母朊病毒NMC菌株。

表1 酵母朊病毒[GPSI+]与[PSI+]对抗朊病毒化合物的敏感性比较Table 1 Comparison of the sensitivities to anti-prion compounds between yeast prions [GPSI+] and [PSI+]
为了深入研究菲啶对酵母朊病毒的治愈作用,我们统计了0.2 mmol/L菲啶作用于酵母菌株NGMC不同时间后的表型分布。如图1所示,阳性对照5 mmol/L盐酸胍作用于白色的[GPSI+]细胞2 d后红色菌落逐渐增多,作用5 d后几乎全为红色菌落;0.2 mmol/L菲啶作用于[GPSI+]细胞3 d天后出现红色菌落,作用5 d后红色菌落的比例为84.6%,而且在菲啶作用[GPSI+]细胞1~5 d内均出现了一定数量的粉色菌落,这与Bach的研究发现基本一致[5]。从表型上看,菲啶作用后的红色菌落初步被认为是治愈的[Gpsi-],白色菌落为未治愈的[GPSI+],粉色菌落可能是一种中间表型,但其细胞内的朊病毒是否聚集或聚集体的大小需要进一步的定量分析。
2.2 细胞水平菲啶对酵母朊病毒[GPSI+]治愈作用的定量分析
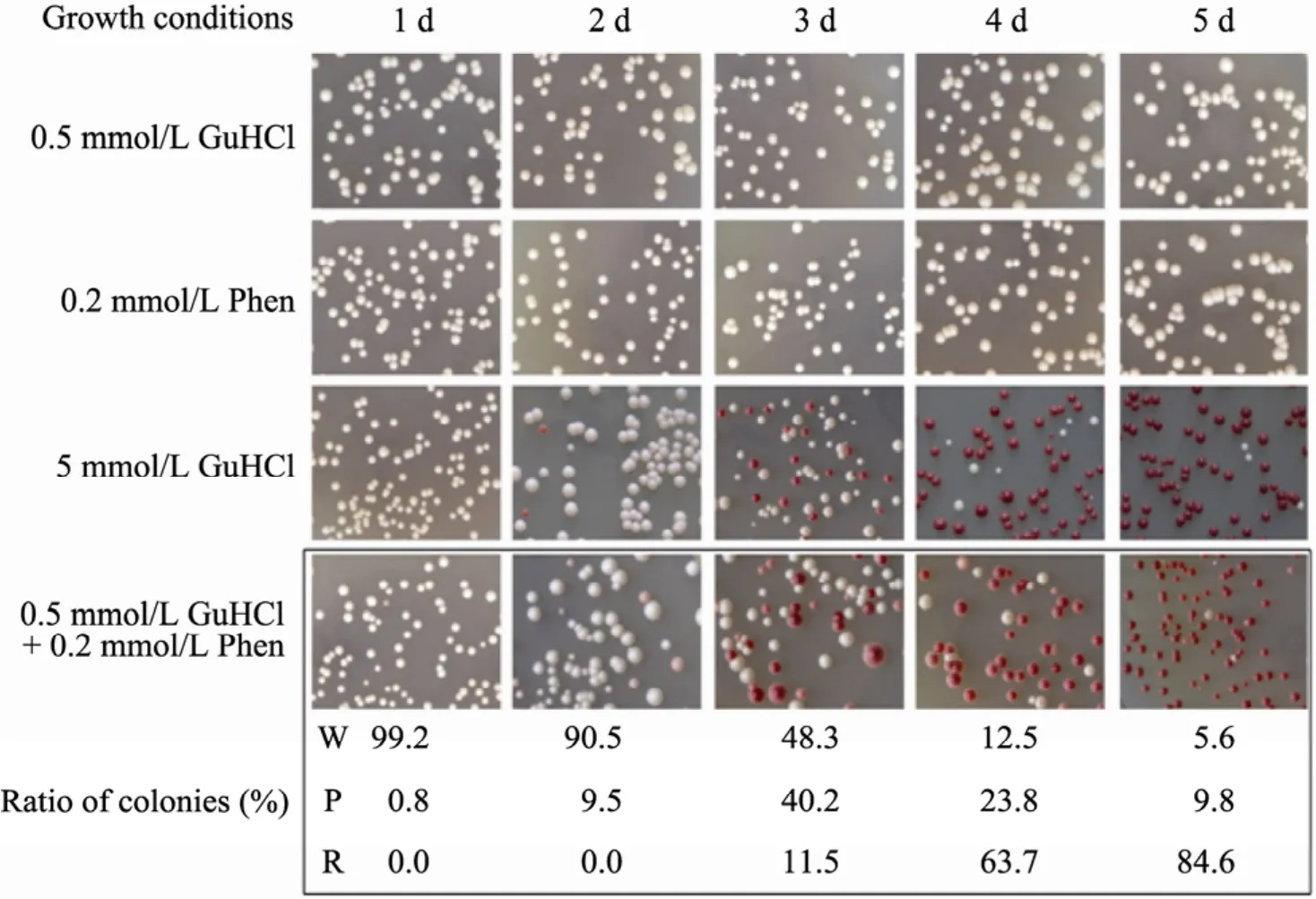
图1 酵母朊病毒[GPSI+]在菲啶作用不同时间后的表型变化Fig. 1 Phenotype changes of [GPSI+] cells after treated by phenanthridine synergized with 0.5 mmol/L GuHCl for 1−5 days. W, P, R represent white, pink and red colonies, respectively.
插入GFP荧光标签的[GPSI+]酵母细胞的朊病毒 (NGMC) 由于聚集而呈现明亮的荧光焦点(Foci),而[Gpsi-]细胞朊病毒是可溶的,因而其荧光呈发散状态。如图2A所示,0.2 mmol/L菲啶与0.5 mmol/L盐酸胍协同作用酵母朊病毒1 d、3 d、5 d后的白色菌落呈现明亮的荧光焦点,说明其朊病毒为聚集状态,而3 d、5 d后的红色菌落其荧光呈发散状态,说明其朊病毒为可溶状态。另外,对照5 mmol/L盐酸胍作用酵母朊病毒[GPSI+]后白色和红色菌落的Foci实验观察结果与菲啶基本一致。为了进一步定量化红色和白色菌落酵母细胞中朊病毒的聚集状态,我们采用了更为精确的荧光漂白后恢复技术 (FRAP) 检测菲啶作用后[GPSI+]细胞中朊病毒NGMC的聚集体大小。如图2B所示,[Gpsi-]细胞的NGMC被漂白后,相对荧光强度迅速上升,大约6 s后趋于平稳,说明[Gpsi-]细胞的 NGMC流动速率快,分子量小;而[GPSI+]细胞的NGMC被漂白后,相对荧光强度升高缓慢,而且在20 s内一直呈上升趋势,说明其NGMC流动速率慢,分子量大。菲啶作用[GPSI+]细胞1 d、3 d后的白色菌落的荧光蛋白光漂白恢复曲线趋势与对照的[GPSI+]细胞类似,而且在20 s内达到的最大相对荧光强度值无显著差异,表明菲啶作用[GPSI+]细胞 1 d、3 d后形成的白色菌落内与对照的[GPSI+]细胞相比较具有大小相当的NGMC聚集体,这说明菲啶作用1 d、3 d后的白色菌落是没有被治愈的[GPSI+]。另一方面,菲啶作用[GPSI+]细胞后3 d、5 d后的红色菌落的NGMC分子量大小与[Gpsi-]细胞的 NGMC分子量大小基本相同,说明这些菌落为被菲啶治愈的[Gpsi-]细胞。值得注意的是,菲啶作用[GPSI+]细胞5 d后的白色菌落的NGMC分子量略小于[GPSI+]细胞,但其朊病毒NGMC仍为聚集状态 (图2A),说明其虽然为[GPSI+]细胞,但其朊病毒聚集体可能发生了轻微的解聚。
2.3 蛋白水平菲啶对酵母朊病毒[GPSI+]治愈作用的定量分析

图2 荧光焦点法和荧光漂白后恢复法定量分析菲啶对酵母朊病毒[GPSI+]的治愈作用Fig. 2 Quantitative analysis of curing effects of phenanthridine on yeast prion [GPSI+] cells with fluorescence foci and FRAP assays. W, R represent white and red phenotype cells respectively. (A) Fluorescence foci assay. (B) Fluorescence redistribution after photobleaching.
为了更为全面地分析菲啶对酵母朊病毒的作用效果,我们在蛋白水平利用半变性琼脂糖凝胶电泳 (SDD-AGE) 结合 Western blotting技术分析化合物作用后[GPSI+]细胞内的朊病毒NGMC聚集体大小。传统的SDS-PAGE只能定性检测朊病毒能否发生聚集,而SDD-AGE利用NGMC聚集体大小。传统的SDS-PAGE只能定性检测朊病毒能否发生聚集,而SDD-AGE利用琼脂糖凝胶的疏松网状结构及朊病毒聚集体独具的 SDS抗性的特点,可以检测朊病毒聚集体的大小[12,17]。如图3所示,[GPSI+]细胞的朊病毒NGMC主要以大分子聚集体形式存在,少量的以可溶的小分子单体形式存在,而[Gpsi-]细胞的朊病毒NGMC以可溶的小分子单体形式存在。菲啶作用[GPSI+]细胞1 d、3 d后的白色菌落的NGMC分子量与[GPSI+]细胞的基本相似 (图 3A),说明这些白色菌落是没有被治愈的[GPSI+];菲啶作用3 d、5 d后的红色菌落的NGMC以单体形式存在,与[Gpsi-]细胞内 NGMC大小基本相同(图3A),说明这些菌落已为治愈的[Gpsi-]细胞。另一方面,菲啶作用后5 d的白色菌落的NGMC仍主要为聚集状态,但分子量较大的NGMC聚集体发生了微弱的解聚,并且可溶的NGMC量比[GPSI+]细胞有少量的增加 (图3A),这与前面FRAP实验所示结果也是一致的 (图2B)。此外,对比图3A和3B发现,盐酸胍作用[GPSI+]细胞后形成的红、白菌落中NGMC大小的变化与菲啶作用后的结果是基本一致的。
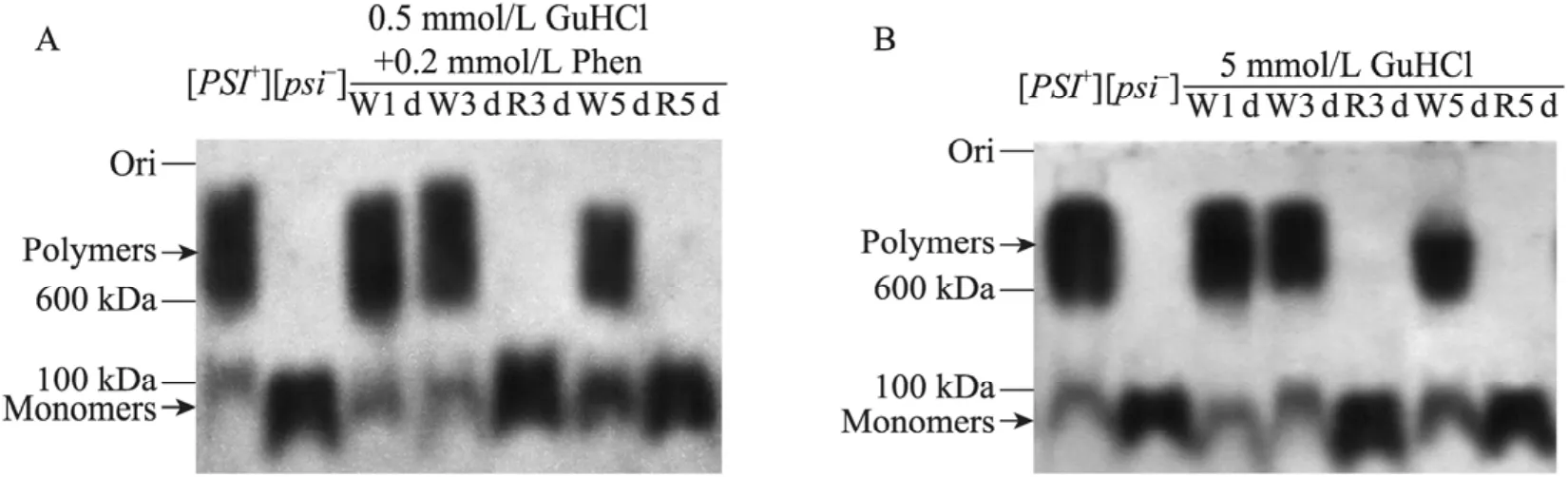
图3 SDD-AGE/Western blotting定量分析菲啶对酵母朊病毒[GPSI+]的治愈作用Fig. 3 Quantitative analysis of curing effects of phenanthridine on yeast prion [GPSI+] cells with SDD-AGE/Western blotting. The migration of molecular size standards (in kilodaltons) is shown on the left. W, R represent white and red phenotype cells, respectively. (A) Phenanthridine. (B) Guanidine hydrochloride.
2.4 菲啶作用后形成粉色中间表型酵母细胞中朊病毒聚集状态的定量分析
综上所述,细胞和蛋白水平的定量分析结果显示,菲啶作用[GPSI+]酵母细胞1 d、3 d、5 d后的白色菌落为非治愈状态的[GPSI+]细胞,而3 d、5 d后的红色菌落为被治愈的[Gpsi-]细胞。利用相同的定量研究方法,我们进一步分析了菲啶作用后形成的粉色中间表型酵母细胞中朊病毒NGMC的聚集状态。如图4A所示,荧光焦点实验分析发现菲啶作用[GPSI+]细胞1 d、2 d后的粉色菌落中的NGMC呈聚集状态,3 d、4 d、5 d后的粉色菌落中的NGMC呈可溶状态。FRAP结果显示菲啶作用[GPSI+]细胞5 d后的粉色菌落的NGMC大小与[Gpsi-]细胞的NGMC大小相当,1 d后的粉色菌落的NGMC大小与[GPSI+]细胞的NGMC大小相似,而2 d、3 d、4 d后的粉色菌落的NGMC大小介于两种对照细胞之间 (图4B)。另一方面,SDD-AGE/Western blotting结果表明菲啶作用[GPSI+]细胞3 d、4 d、5 d后的粉色菌落的NGMC主要以单体形式存在;而1 d后的粉色菌落的NGMC主要以聚集体形式存在,与[GPSI+]细胞的朊病毒聚集体大小相似;2 d后的粉色菌落的NGMC同样主要以聚集体形式存在,但其分子量略低于[GPSI+]细胞中的NGMC聚集体,并且可溶的单体NGMC含量明显增多 (图 4C),这与 FRAP的结果是基本一致的。
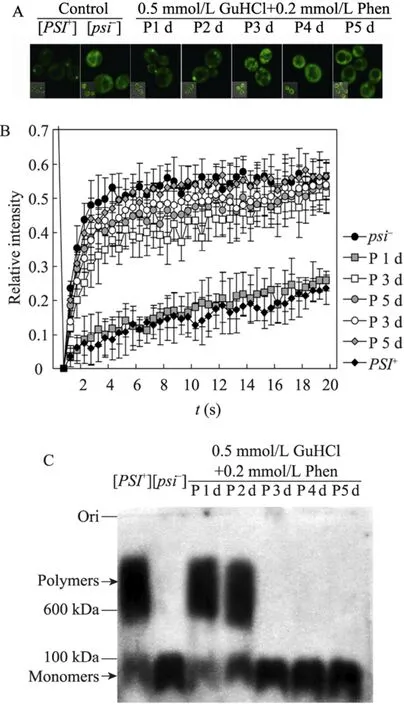
图4 细胞和蛋白水平定量分析菲啶作用酵母朊病毒[GPSI+]后粉色表型的朊病毒聚集状态Fig. 4 Quantitative analysis of the aggregation status of Sup35p in phenanthridine induced pink (P) phenotype cells at the cellular and protein levels. (A) Fluorescence foci. (B) Fluorescene redistribution after photobleaching. (C) SDD-AGE/Western blotting.
3 讨论
我们前期研究结果显示,5 mmol/L盐酸胍对[GPSI+] (NGMC) 菌株的表型治愈效果与 Bach等使用的野生型酵母[PSI+] (NMC) 菌株的效果是相似的[20]。本研究进一步发现抗朊病毒化合物菲啶对酵母朊病毒[GPSI+]的表型治愈作用类似于野生型酵母[PSI+]菌株 (NMC),这暗示着可以用酵母朊病毒[GPSI+]菌株替代野生型[PSI+]菌株以便利用新的方法实现细胞及蛋白蛋白水平的定量分析。另外,Bach等研究发现,菲啶单独存在只能治愈药物敏感型酵母朊病毒菌株([PSI+]的强度较弱) 而不能治愈野生型菌株[5]。而药物敏感型菌株生长非常缓慢,难于进行细胞及蛋白水平的定量研究。为此,我们借鉴了Bach等提出的使用增效剂盐酸胍的方法提高本实验体系中[GPSI+]菌株对药物的灵敏度[5]。
借助本实验室构建独有的酵母朊病毒[GPSI+]细胞模型,我们采用 SDD-AGE/Western blotting技术和 FRAP技术在蛋白和细胞水平定量分析了菲啶对酵母朊病毒的治愈作用,从而弥补了此前研究对菲啶的抗酵母朊病毒效果缺乏蛋白和细胞水平定量分析的不足和对粉色表型没有精确定量分析的缺陷。研究结果表明,菲啶作用酵母朊病毒[GPSI+]细胞1 d、2 d后出现的粉色菌落中朊病毒的聚集状态与[GPSI+]相似,而3~5 d后出现的粉色菌落中朊病毒的状态与[Gpsi-]相似,呈现出密切的时间相关性。结合前期研究中提出的酵母朊病毒的治愈是通过细胞分裂降低细胞质中朊病毒“种子”的假说[12-14],这暗示着菲啶完全治愈[PSI+]需要更多的细胞分裂周期。另外,值得注意的是,酵母朊病毒的颜色表型与其在细胞中的聚集状态虽然总体上一致,但同时存在着细微的区别。这暗示着在统计菲啶对朊病毒的治愈率时,不能简单地认为红色菌落的比例代表了菲啶的治愈率,而应根据不同菌落的系统检测结果来具体分析,因而菲啶作用酵母朊病毒[GPSI+] 1~5 d的治愈率分别为0%、0%、51.7%、87.5%和94.4% (图1)。此外,本研究结果也显示蛋白水平采用的 SDD-AGE/ Western blotting技术和细胞水平采用的FRAP技术在定量研究抗朊病毒候选药物的作用效果的重要性和可行性,极大地丰富了Bach等提出的基于酵母细胞的抗朊病毒药物筛选模型的次级筛选分析方法。
[1] Prusiner SB. Biology of prion diseases. J Acquir Immune Defic Syndr, 1993, 6(6): 663−665.
[2] Ghaemmaghami S, May BCH, Renslo AR, et al. Discovery of 2-aminothiazoles as potent antiprion compounds. J Virol, 2010, 84(7): 3408−3412.
[3] Sim VL, Caughey B. Recent advances in prion chemotherapeutics. Infect Disord Drug Targets, 2009, 9(1): 81−91.
[4] Trevitt CR, Collinge J. A systematic review of prion therapeutics in experimental models. Brain, 2006, 129(9): 2241−2265.
[5] Bach S, Talarek N, Andrieu T, et al. Isolation of drugs active against mammalian prions using a yeast-based screening assay. Nat Biotechnol, 2003, 21(9): 1075−1081.
[6] Bach S, Tribouillard D, Talarek N, et al. A yeast-based assay to isolate drugs active against mammalian prions. Methods, 2006, 39(1): 72−77.
[7] Tribouillard-Tanvier D, Béringue V, Desban N, et al. Antihypertensive drug guanabenz is active in vivo against both yeast and mammalian prions. PLoS One, 2008, 3(4): e1981.
[8] Zhong ZW, Song YT. Recent advances of research on Antiprion candidate drugs. Chin J Zoonoses, 2011, 27(9): 828−835.钟正伟, 宋有涛. 朊病毒治疗候选药物的研究进展. 中国人兽共患病学报, 2011, 27(9): 828−835.
[9] Song YT, Wu XY, He XR. Development of the model for antiprion drug screening. J Liaoning Univ Nat Sci: Nat Sci Edi, 2009, 37(4): 289−293.宋有涛, 吴宪远, 何星蓉. 抗朊病毒药物筛选模型的研究进展. 辽宁大学学报: 自然科学版, 2009, 37(4): 289−293.
[10] Ishikawa T. Recent advances of research on the [PSI+] prion in Saccharomyces cerevisiae. Mycoscience, 2008, 49(4): 221−228.
[11] Tuite MF, Mundy CR, Cox BS. Agents that cause a high frequency of genetic change from [PSI+] to [psi-] in Saccharomy cescerevisiae. Genetics, 1981, 98(4): 691−711.
[12] Kryndushkin DS, Alexandrov IM, Ter-Avanesyan MD, et al. Yeast [PSI+] prion aggregates are formed by small Sup35 polymers fragmented by Hsp104. J Biol Chem, 2003, 278(49): 49636−49643.
[13] Song YT, Wu YX, Jung GM, et al. Role for Hsp70 chaperone in Saccharomyces cerevisiae prion seed replication. Eukaryot Cell, 2005, 4(2): 289−297.
[14] Song Y, Lan WJ, Wu XY, et al. Quantitative effects of magnesium chloride stress on aggregation of Sup35p in [psi-] yeast cells. Protein Pept Lett, 2010, 17(12): 1489−1494.
[15] Wu YX, Greene LE, Masison DC, et al. Curing of yeast [PSI+] prion by guanidine inactivation of Hsp104 does not require cell division. Proc Natl Acad Sci USA, 2005, 102(36): 12789−12794.
[16] Wu YX, Masison DC, Eisenberg E, et al. Application of photobleaching for measuring diffusion of prion proteins in cytosol of yeast cells. Methods, 2006, 39(1): 43−49.
[17] Halfmann R, Lindquist SL. Screening for amyloid aggregation by semi-denaturing detergent-agarose gel electrophoresis. J Vis Exp, 2008, 17(1): 1−4.
[18] Greene LE, Park YN, Masison DC, et al. Application of GFP-labeling to study prions in yeast. Protein Pept Lett, 2009, 16(6): 635−641.
[19] Wickner RB. [URE3] as an altered URE2 protein: evidence for a prion analog in Saccharomyces cerevisiae. Science, 1994, 264(5158): 566−569.
[20] Song YT, Song Y, Zhong ZW, et al. Illuminating precise quantification in yeast-based model for antiprion compounds screening. 2010 First International Conference on Cellular, Molecular Biology, Biophysics and Bioengineering. Qiqihar: Institute of Electrical and Electronics Engineers Press, 2010, 1: 296−299.
猜你喜欢
食品工业科技(2022年23期)2022-12-06
大自然探索(2022年5期)2022-07-11
VOGUE服饰与美容(2022年5期)2022-05-01
河南工业大学学报(自然科学版)(2021年6期)2022-01-26
化工科技(2021年5期)2021-11-24
今日农业(2021年10期)2021-07-28
世界科学(2020年11期)2020-11-24
化工学报(2020年10期)2020-10-27
中国兽医杂志(2018年4期)2018-01-25
医药前沿(2018年33期)2018-01-16
