
东方田鼠抗日本血吸虫病CD74基因的差异表达及生物信息学分析
2012-09-20 03:29胡君健郑学琴
湖南理工学院学报(自然科学版) 2012年2期
向 阳 , 胡君健, 郑学琴
(1. 湖南理工学院 化学化工学院, 湖南 岳阳 414006; 2. 湖南省血吸虫病防治研究所, 湖南 岳阳 414006)
东方田鼠抗日本血吸虫病CD74基因的差异表达及生物信息学分析
向 阳1, 胡君健2, 郑学琴1
(1. 湖南理工学院 化学化工学院, 湖南 岳阳 414006; 2. 湖南省血吸虫病防治研究所, 湖南 岳阳 414006)
利用日本血吸虫尾蚴感染东方田鼠, 提取感染前和感染后10d 和15d东方田鼠肝脏组织总RNA; 利用大鼠CD74基因探针, 经Rorthern杂交分析东方田鼠感染日本血吸虫前及感染血吸虫10 d、15 d后肝脏组织CD74的差异表达情况;同时, 采用生物信息学分析大鼠CD74基因的cDNA序列及其编码的氨基酸序列以及CD74蛋白的结构域. 结果显示: 东方田鼠感染日本血吸虫感染血吸虫10 d、15 d后肝脏组织CD74的表达水平比感染前显著升高; 大鼠CD74基因的cDNA序列全长1220bp, 编码216个氨基酸残基; 大鼠CD74蛋白含一个MHC2相互作用超家族结构域和一个MHCassoc_trimer超家族结构域.
东方田鼠; 日本血吸虫;CD74; 差异表达; 生物信息学分析
1 材料和方法
1.1 材料
1.1.1 动物
东方田鼠为室内繁殖的湖南洞庭湖种, 由湖南省血吸虫病防治研究所提供.
1.1.2 日本血吸虫尾蚴
由湖南省血吸虫病防治研究所提供新鲜逸出的日本血吸虫尾蚴, 进行人工定量感染试验.
1.2 方法
1.2.1 组织样品收集和RNA 提取
用1000条日本血吸虫尾蚴感染东方田鼠, 于感染10d 和15d后取东方田鼠肝脏组织及阳性对照, 提取总RNA见文献[18].
1.2.2 Northern杂交实验
探针制备及Northern杂交见文献[19]. 本实验芯片杂交实验操作如下: 分别利用大鼠CD74基因探针与未感染的东方田鼠和感染10d和15天后的东方田鼠肝组织样品mRNA杂交.
1.2.3 生物信息学分析
美国国家生物信息中心NCBI EST数据库(http://www.ncbi.nlm.nih.gov/)和Unigene数据库(http://www.ncbi.nlm. nih.gov/Unigene). 阅读框架的识别: 利用ExPASy服务器中的 Translate和NCBI中的ORF finder. 结构域分析: 利用NCBI中BLAST(http://blast.ncbi.nlm.nih.gov/Blast.cgi)的protein blast.
2 结果
2.1 感染日本血吸虫前后东方田鼠肝脏组织RNA抽提
抽提东方田鼠感染日本血吸虫前及感染10 d、15 d后肝组织总RNA. 按《分子克隆》说明抽提, 每条泳道上上样mRNA约3 μg (图1).
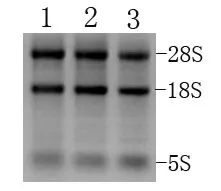
图1 东方田鼠感染日本血吸虫前后肝脏组织总RNA抽提结果
2.2 东方田鼠正常对照组与感染日本血吸虫10 d、15 d后肝组织CD74的差异表达
利用大鼠CD74基因探针, 经Rorthern杂交分析, 东方田鼠感染日本血吸虫10 d、15 d后肝组织CD74的表达比正常对照组显著上调(图2).
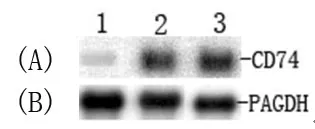
图2 Northern blot分析CD74
2.3 CD74 基因完整阅读框的生物信息学分析
大鼠CD74基因的cDNA 序列全长1220bp, 编码216个氨基酸残基(图3).

图3 大鼠CD74基因的cDNA序列及其编码的氨基酸序列
2.4 CD74 蛋白结构域的生物信息学分析
大鼠CD74蛋白含216个氨基酸残基,含一个MHC2相互作用超家族结构域和一个MHCassoc_trimer超家族结构域(图4).

图4 大鼠CD74蛋白结构域分析
3 讨论
血吸虫病是继疟疾之后的全球第二大寄生虫病, 严重危害人类健康. 中国是血吸虫病的重灾区之一. 我国流行的主要是日本血吸虫病, 全国累计患者达1161.2万人, 受血吸虫病威胁的人口达1.3亿. 东方田鼠具有天然抗日本血吸虫能力, 是日本血吸虫的非适宜性宿主.利用大鼠CD74基因探针与未感染日本血吸虫尾蚴的东方田鼠和感染日本血吸虫尾蚴后第10 d和15d的东方田肝组织mRNA杂交, 感染10 d和15d肝脏内CD74基因的表达显著上调, CD74作为一种跨膜蛋白是巨噬细胞迁移抑制因子( macrophage m igration inh ib itory factor, M IF)的受体分子参与免疫免疫调节, 表明东方田鼠感染日本血吸虫后机体免疫系统得到积极应答. 同时, 本文对大鼠CD74基因进行了生物信息学分析,CD74全长1220bp, 编码216个氨基酸残基, 含含一个MHC2相互作用超家族结构域和一个MHCassoc_trimer超家族结构域. 本研究为进一步研究CD74在东方田鼠抗日本血吸虫病防治中的作用奠定了一定基础.
[1] He Y, Luo X, Zhang X, et al.Immunological characteristics of natural resistance in Microtus fortis to infection with Schistosoma japonicum[J]. Chin Med J.1999, 112(7): 649~654
[2] 邹国军, 胡维新. 东方田鼠天然抗日本血吸虫病机制研究进展[J]. 中国地方病学杂志, 2006, 25 (2): 227~228
[3] 俞远京, 胡维新, 王 勇, 等. 野生东方田鼠实验动物化及种质资源保护的初步研究[J]. 实验动物科学与管理, 2003, 20: 13~17
[4] 李 浩, 何艳燕, 林邦发, 等. 东方田鼠重复感染日本血吸虫试验初步研究[J]. 中国兽医寄生虫病, 2001, 9(3): 11~17
[5] 刘杰生, 容寿铭, 陈佩玑, 等. 几种鼠类人工感染日本血吸虫的实验观察[J]. 中国血吸虫病防治杂志, 1992,4( 6): 350~351
[6] 刘金明, 傅志强, 李 浩, 等. 东方田鼠血清体外杀伤日本血吸虫童虫效果的初步观察[J]. 中国人兽共患病杂志, 2002, 18(2): 82~84
[7] 蒋守富, 邱倩雯, 何艳燕, 等. 东方田鼠血清蛋白组分体外杀伤日本血吸虫童虫作用的研究[J]. 中国血吸虫病防治杂志, 2008, 20(4):260~264
[8] 蒋守富, 魏梅雄, 林矫矫, 等. 东方田鼠IgG3 抗体抗血吸虫病作用研究[J]. 中国寄生虫学与寄生虫病杂志, 2008, 26 (10): 34~36
[9] 阎玉涛, 刘述先, 宋光承, 等. 东方田鼠天然抗体相关的日本血吸虫抗原基因筛选和克隆[J]. 中国寄生虫学与寄生虫病杂志, 2001, 19(3): 153~156
[10] 袁忠英, 沈玉娟, 曹建平, 等. 东方田鼠血清免疫筛选日本血吸虫童虫cDNA文库及新基因分析[J]. 中国血吸虫病防治杂志, 2008, 20(4): 552-556
[11] 孙 毅, 孙 焕, 贾人初, 等. 东方田鼠抗日本血吸虫抗性相关靶基因筛选[J]. 中国血吸虫病防治杂志, 2008, 20(1): 26~31
[12] 秦志强, 胡维新, 邬国军, 等. 东方田鼠骨髓基因池的构建及抗日本血吸虫抗性相关基因的筛选[J]. 生命科学研究, 2004, 8(4): 333~338
[13] 孙 军, 林矫矫, 程国锋, 等. 利用基因表达谱芯片研究东方田鼠和小鼠感染日本血吸虫前后基因的差异性表达[J]. 北京大学学报(自然科学版) , 2004, 40(4): 532~537
[14] K leemann R, H ausserA, Ge ige rG, et al.Intracellular action o f the cytokine M IF to m odu late AP1 activity and the cell cycle through Jab1[J].Nature, 2000, 408 ( 6809): 211~216
[15] M cC lellandM, Zhao LJ, Carskadon S, et al.Expression o f CD74, the recepto r form acrophage mig- ration inhibitory factor, innonsma ll cell lung cancer[J] . Am J Patho, 2009, 174( 2): 638~646
[16] Cho YS, Jones BF, Verm e ire JJ, et al.Structural and functiona l character ization of a secreted hookworm m acrophage m igration inh ib itory factor (M IF) that interacts w ith the humanM IF receptor CD74[ J] . J Biol Chem, 2007, 282 ( 32): 23447~23456
[17] Dobson SE, Augustijn KD, B rann igan JA, et al.The crysta l structures o fm acrophage migration inh- ibitory factor from Plasmodium falciparum and Plasmodium berghei[J] . Protein Sci, 2009, 18( 12) : 2578~2591
[18] Y. XIANG, D. S. NIE, Q. J. ZHANG,et al..Cloning, characterization and identification of Rcet1-v1 and Rcet1-v2, two novel splice variants of mouse Rcet1 related to Cres subgroup of family 2 cystatins[J]. DNA sequence the journal of DNA sequencing and mapping, 2008,19(1): 13~19
[19] Yang XIANG, Dong-Song NIE, Jian WANG, et al..Cloning, Characterization and Primary Function Study of a Novel Gene, Cymg1, Related to Family 2 Cystatins[J]. Acta Biochimica et Biophysica Sinica, 2005, 37(1): 11~18
Different Expression and Bioinformatic Analysis of Microtus Fortis in Anti- Schistosomiasis Japonica CD74 Gene
XIANG Yang1, HU Jun-jian2, ZHENG Xue-qin1
(1. College of Chemistry and Chemical Engineering, Hunan Institute of Science and Technology, Yueyang 414006, China; 2. Hunan Provincial Institute of Schistosomiasis Control, Yueyang 414006, China )
The total RNA was extracted from microtus fortis liver tissue which was infected of the schistosoma japonicum cercariae before infection and after infection 10d and 15d. Using rattus norvegicusCD74gene probe to hybridize analysis ofCD74difference expression in the microtus fortis liver tissues which were infected with Schistosoma japonicum before infection and after infection 10 d, 15 d. At the same time, bioinformatics analysis of the cDNA sequence and encoded amino acid sequence of the rattus norvegicusCD74gene and CD74 protein structural domains. The results showed: after microtus fortis infected with schistosoma japonicum 10 d and 15 d the liver tissue ofCD74expression levels significantly higher than pre-infection; rattus norvegicusCD74cDNA sequence of a total length is 1220bp and encode 216 amino acid residues; rattus norvegicus CD74 protein containing a MHC2 interaction domain of the superfamily and a MHCassoc_trimer superfamily domain.
microtus fortis; schistosoma japonicum;CD74; differences in expression; bioinformatic analysis
Q752
A
1672-5298(2012)02-0044-03
血吸虫病是继疟疾之后的全球第二大寄生虫病, 严重危害人类健康. 全球至少有76个国家和地区受到危害, 约有6亿人受到威胁, 2亿人被感染, 而中国是血吸虫病的重灾区之一. 我国流行的主要是日本血吸虫病, 全国累计患者达1161.2万人, 受血吸虫病威胁的人口达1.3亿. 我国血吸虫病主要分布在江苏、湖南、湖北、四川和云南等省, 洞庭湖区为全国重灾区之一, 感染人群占全国的1/4. 东方田鼠 (Microtus fortis) 是日本血吸虫的非适宜宿主, 具有天然完全抗日本血吸虫病的特性[1]. 东方田鼠在我国的分布主要集中在长江流域, 特别是在日本血吸虫病流行的洞庭湖湖洲, 是该地域的一个优势鼠种[2]. 日本血吸虫感染东方田鼠后第12天开始, 虫体生长发育停滞, 第20~28天虫体在体内全部消亡[3], 日本血吸虫在东方田鼠体内的消亡部位主要是肝脏[4]. 实验表明: 鼠类中, 黄毛鼠、板齿鼠、海南屋顶鼠、黄胸鼠、大鼠对日本血吸虫的易感性由高到低[5], 大鼠为日本血吸虫非适宜性宿主. 这揭示了不同鼠种对日本血吸虫有不同的抵抗能力. 至今, 几乎所有的血吸虫感染动物模型都是阳性模型, 而东方田鼠感染血吸虫模型却是一种阴性模型. 因此, 利用东方田鼠天然抗日本血吸虫感染的动物模型, 我国科学家对东方田鼠的体液免疫和细胞免疫[6~8]、天然抗体相关抗原基因[9~11]、抗病基因的筛选[12]、利用基因表达谱芯片筛选东方田鼠感染日本血吸虫前后的差异基因[13]及功能研究等做了大量的研究工作. CD74作为一种跨膜蛋白是巨噬细胞迁移抑制因子( macrophage m igration inh ib itory factor,M IF)的受体分子, 与多种人类自身疾病的发生和寄生虫的感染相关[14], 可在57 种肿瘤细胞中检测到CD74基因的高表达, 并且是以CD74-M IF 复合体的方式存在[15]; CD74还可与寄生虫来源的M IF 相互作用, 其结合机制以及相互作用后在调节宿主细胞效应方面与宿主M IF均有不同[16,17]. 本文比较了东方田鼠感染日本血吸虫前后CD74的差异表达及对CD74进行了生物信息学分析, 为进一步研究CD74在东方田鼠抗日本血吸虫病的机理打下了一定基础.
2012-04-10
湖南省教育厅重点项目(09A035); 湖南省卫生厅一般项目(B2009-102)
向 阳(1968- ), 男, 湖南怀化人, 博士, 湖南理工学院化学化工学院教授. 主要研究方向: 分子生物学
猜你喜欢
作文大王·低年级(2021年11期)2021-12-22
小天使·一年级语数英综合(2021年2期)2021-05-08
现代畜牧科技(2021年2期)2021-03-19
热带病与寄生虫学(2021年2期)2021-02-28
小学生(看图说画)(2020年4期)2020-04-01
中国感染与化疗杂志(2020年1期)2020-01-12
家庭用药(2018年3期)2018-03-23
湖南畜牧兽医(2016年3期)2016-06-05
兽医导刊(2016年6期)2016-05-17
河南畜牧兽医(2015年13期)2015-11-28
