
短花针茅叶片解剖结构及与气候因子的关系
2013-04-10 06:13康萨如拉牛建明张庆陈丽萍
草业学报 2013年1期
康萨如拉,牛建明,2*,张庆,陈丽萍,3
(1.内蒙古大学生命科学学院,内蒙古 呼和浩特010021;2.中美生态、能源及可持续性科学研究中心,内蒙古 呼和浩特010021;3.南开大学生命科学学院,天津300071)
植物个体生长于自然环境中,自然受到干旱、盐碱、低温(冷害、冻害)、高温等气候因子的影响,然而植物对生存的不良环境具有特定的适应性和抵抗力[1-3]。植物对逆境适应与响应过程中有形态结构、解剖结构以及生理生化上的变化[4,5]。生长在不同生境中的植物,常表现出不同的形态结构;同种植物因生境的显著差异,其形态和解剖结构也会表现出明显的趋异现象[6,7],这与陈庆诚等[8]的多途径适应观点一致。植物形态结构能反映气候因子对植物的影响以及植物自身对环境的适应,这通常被认为是植物对特殊环境的适应反应[9,10]。
植物叶片是反映植物对环境适应的最敏感器官,也是植物生命活动进行的主要部位[9]。因此在不同生境条件下,植物叶片的各个结构对其所处的环境有各自的响应特征[11]。暴露在空气中的植物叶片大小、形状及方向性在一定程度上决定着植物和环境之间能量相互转换的效率[12-16]。Hegazy和Elamry[17]认为植物叶片形态差异会共同表现在植物叶片大小和形状上,而恰恰这种特征保证了植物和环境之间物质和能量的相互转换,是植物在干旱、半干旱环境中生存的重要条件。还有些研究指出,对于不同地段的植物群落,其叶片的类型、形状、结构是所处环境最独特的标志,并且认为物种的分化不仅表现在其叶片的外部形态结构,在叶片解剖结构及化学成分上也有极大的不同[18-20]。因此,研究植物叶片形态及解剖结构的分化特征,对植物适应性和抗性生理研究具有重要意义。
短花针茅(Stipabreviflora)系禾本科(Gramineae)针茅族(Stipeae)针茅属(Stipa)须芒组(Sect.Barbatae)多年生丛生型旱生草本植物,是亚洲中部暖温型荒漠草原的主要建群种,也常在某些典型草原群落及草原化荒漠群落中成为伴生成分[21]。短花针茅草原是亚洲中部草原亚区荒漠草原带气候偏暖地区的主要草原类型,它的分布从我国黄土丘陵区西北部起,向北越过阴山山地到内蒙古高原的南部地区,其南界大体上达到甘肃的兰州、会宁、宁夏的固原、陇东的环县,陕北的靖边、榆林、绥德以及晋西北的河曲、偏关等地。短花针茅草原分布区的湿润度在0.23~0.47,年降水量为279(化德)~434(准格尔)mm,年均温为2.1(化德)~7.4℃(准格尔),兰州可达9.1℃[22]。因此,短花针茅适于研究植物适应性及抗性生理[23,24]。本研究通过短花针茅叶片解剖结构对气候因子的适应性研究,试图阐明导致短花针茅叶片结构差异的主导环境因子,探究短花针茅适应环境的策略,为植物适应性和抗性生理研究及相关领域提供理论参考。
1 材料与方法
1.1 研究区概况
本研究设置野外调查样地共8个,分布于内蒙古准格尔旗皇甫川、鄂托克旗三北羊场、鄂托克旗东深井、达茂旗巴音敖包苏木、达茂旗百灵庙、达茂旗石宝乡、苏尼特右旗都仁乌力吉苏木(图1)。其中,在苏尼特右旗都仁乌力吉苏木选取2个不同生境下的样地,其地理位置介于北纬39.24°~42.48°、东经107.34°~113.33°。样地的植被类型包括温性典型草原、温性荒漠草原、暖温性典型草原、暖温性荒漠草原和草原化荒漠,详情见表1[25]。

图1 样地分布Fig.1 The distribution of observation sites
1.2 研究方法
1.2.1 取样 2007年8月,在研究区各样地随机剪取短花针茅(依据内蒙古植物志[22]及内蒙古大学植物标本馆中短花针茅标本对所取样品进行鉴定)植株相同高度的成熟、健康、向阳、无虫害的基生叶3.0~3.5 cm,重复取样25~30株,迅速放入福尔马林-醋酸-酒精固定液(formalin-acetic acid-alcohol stationary liquid,FAA)中固定,带回实验室4℃保存备用。
1.2.2 制作石蜡切片及成像 将FAA固定液中固定的短花针茅叶片(每个样地随机选择5~6株),通过脱水、透明、渗蜡、石蜡包埋、修蜡、切片、粘片(每片粘10~20横切面,如图2,每株重复3次)、烘片、脱蜡、双重染色及封片等步骤,制作短花针茅叶片的石蜡切片[26,27]。
在OLYMPUS CX41显微镜下分别对切片进行4×10倍和10×10倍观察,筛选出染色效果较佳的切片,使用CANON DS16275数码相机以照片与切片100∶1的比例进行拍照与存储,显微照片示例图像见图3。并按JPEG格式存储图像文件。根据测微尺的刻度确定图像的长和宽,分别为1 550.24和1 033.49 μm。
1.2.3 图像分析及数据获取 首先将JPEG图像转化为PIX图像文件,在PCI Geomatica V 8.2软件下,确定每张图片左下角及右上角的坐标,定义单位为微米(μm)。
在 ArcMap version 9.3下每个样地选取10~15幅染色及成像效果较佳的切片,对其实际结构进行矢量化。该处主要对叶片表皮、角质层、栅栏组织、维管束、主脉导管腔进行了矢量化与拓扑,形成Coverage文件[28]。

图2 短花针茅叶片石蜡切片Fig.2 Paraffin section of S.breviflora

图3 短花针茅显微照片示例(1mm,100∶1)Fig.3 Micro-image of S.breviflora

表1 样地描述[25]Table 1 The description of observation sites

续表1 Continued
在ArcMap version 9.3下对叶片结构的长度、厚度、面积等参数进行测量统计后,确定以下6个指标,并获得每个样地指标的均值。
A:角质层占叶片厚度的比例(%)=角质层厚度/叶片厚度×100
B:维管束占叶面积比例(%)=维管束总面积/叶面积×100
C:栅栏组织占叶片厚度的比例(%)=栅栏组织厚度/叶片厚度×100
D:主导管占主维管束比例(%)=主维管束导管面积/主维管束面积×100
E:表皮细胞密度(%)=表皮细胞个数/统计长度×100
F:表皮毛密度(%)=表皮毛个数/统计长度×100
1.3 样地与气候指标的获得
选取10个气候因子,即≥0℃年积温(AMT0)、≥10℃年积温(AMT10)、最冷月平均气温(CMT)、最热月平均气温(WMT)、年平均气温(MT)、年蒸发量(EVP)、年降水量(P)、夏秋降水量(P_SF)、冬春降水量(P_WS)及无霜期(FRZ_FD)。依据内蒙古气候要素空间分布模型[29],基于调查地点的经度、纬度与海拔,分别获取8个样地的上述10个气候因子指标。
1.4 统计分析
利用Canoco For Windows 4.5软件,对叶片结构数据矩阵(6个指标)与气候数据矩阵(10个指标)进行典范对应分析(canonical correspondence analysis,CCA),阐明叶片结构与气候因子之间的关系,探究影响短花针茅分布及叶片结构分化的主导气候因子。
结合上述CCA分析结果,选取2个主导气候因子年平均气温和年降水量,利用SPSS 16.0,对叶片结构数据矩阵(6个指标)与气候因子(2个指标)之间进行Pearson相关显著性检验,阐明主导气候因子与各个叶片结构的关系,探讨短花针茅适应环境时所采取的策略。
结合8个样地短花针茅的遗传分化[25],对短花针茅叶片结构和遗传结构数据矩阵之间进行Mantel检验,阐明遗传分化在短花针茅叶片结构分化中所起的作用。
2 结果与分析
2.1 短花针茅叶片的结构特征
短花针茅叶片有以下典型的旱生特征(图4)。如叶片横切面完全卷曲成“V”型或椭圆型,表皮细胞排列整齐密集,上表皮有致密的表皮毛,有发达紧密的维管组织及维管束有发达的导管腔,叶片上表皮弯曲的每个凹陷处有数个运动细胞,厚壁组织排列紧密,具有加厚的角质层,有较大的气孔密度等。
统计表明,短花针茅叶片厚度为(122.42±5.28)μm,角质层加厚,为(5.23±0.63)μm,表皮细胞密度为(4.90±0.36)个/μm,维管束总面积占叶面积的比例为(10.52±0.87)%,靠近下表皮的栅栏组织厚度为(38.16±3.29)μm,栅栏组织厚度占叶片厚度的比例为(31.44±1.64)%(表2)。
比较发现,不同样地短花针茅主维管束导管腔占主维管束面积比值的差异最大,其次为表皮毛密度(表2)。而进一步对不同样地间短花针茅叶片结构分化差异显著性检验表明(表3),叶片结构分化具有极显著性差异的有样地8与样地1,2,3,7之间和样地2与样地5,6之间。样地8与样地4,5和6之间的叶片结构分化差异显著。

图4 石蜡切片Fig.4 Parafin section

表2 6个指标统计数值表Table 2 Numerical value of six indices
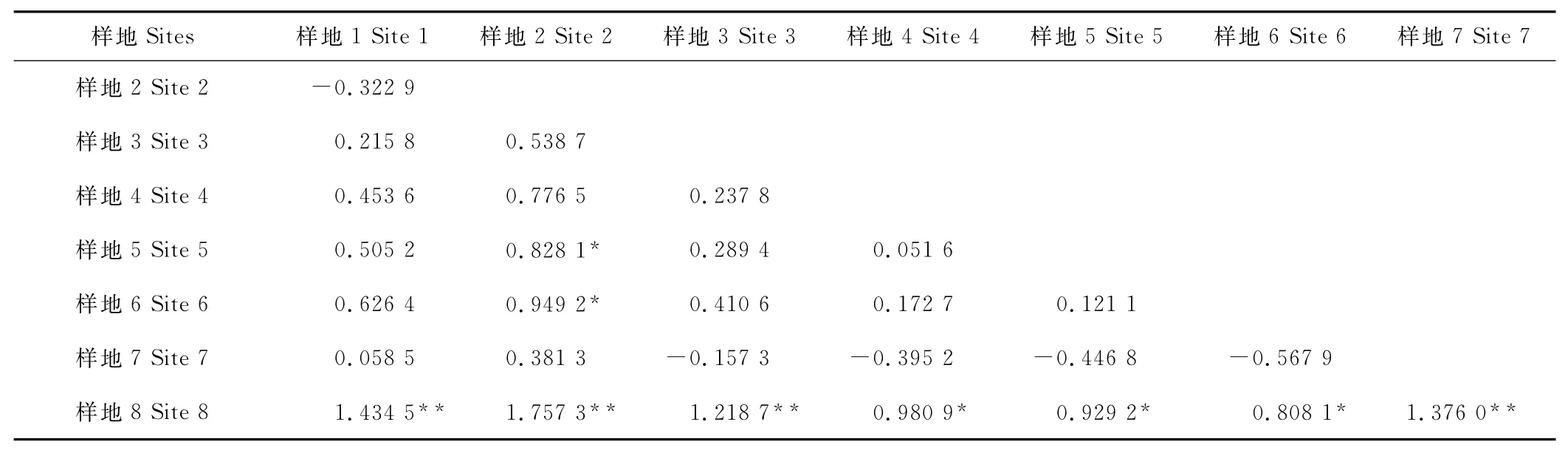
表3 样地之间叶片结构差异显著性检验Table 3 Variable significant analysis of eight sites
2.2 叶片结构与气候因子的关系
第1主导因子为热量因子(83.0%),而水分因子位居第2(13.9%)(图5)。8个样地在第1轴上受AMT0℃、AMT10℃、MT、FRZ_FD、CMT、WMT的影响较大,在第2轴上受P_SF、P、P_WS的影响较大。1,2和6,7,8样地受热量因子的影响较大;3和4,5样地受水分因子的影响较大。1,2与热量因子正相关,而6,7,8三个样地与热量因子负相关;第3样地与水分因子正相关,4,5两个样地与水分因子负相关。

图5 CCA分析结果Fig.5 The result of CCA
通过对信息量相关性矩阵的分析发现(表4),在热量因子当中年平均气温对8个样地短花针茅的影响最大(0.763),而≥0℃年积温与≥10℃年积温位居第2(0.762 8和0.759 1);结合排序结果,无霜期因子最接近第1轴,说明无霜期与热量的相关性非常显著。水分因子当中年平均降水量的影响最大(0.803 3),夏秋降水量居第2位;年降水量与夏秋降水量的相关性比与冬春降水量的相关性大,说明一年中夏秋降水量对8个样地短花针茅生长过程中的影响比冬春降水量的影响大。
Pearson显著性检验结果表明(表5),维管束占叶面积的比值和主维管束导管占主维管束面积的比值与热量因子呈显著正相关,而角质层厚度占叶片厚度的比值、栅栏组织厚度占叶片厚度的比值、表皮细胞密度、表皮毛密度与热量因子的相关性不显著。
与水分因子具有显著相关性的指标为维管束占叶面积的比值,其他指标均不显著。不过,尽管角质层厚度/叶片厚度、栅栏组织厚度/叶片厚度、表皮细胞密度与水分因子之间的相关性不显著,但具有负相关特征,即随着年降水量减小,角质层厚度增大,表皮细胞密度增加,栅栏组织厚度增大,符合植物抗旱性的特点。
2.3 叶片结构与遗传特征的关系
对短花针茅叶片解剖结构与遗传结构之间的Mantel检验结果表明(图6),短花针茅叶片解剖结构与遗传特征之间存在相似性,但不显著。

表4 CCA信息量相关性矩阵Table 4 Information table of CCA

表5 Pearson相关显著性检验结果Table 5 Result of Pearson test
3 结论与讨论
3.1 短花针茅叶片具有典型的旱生结构
为了更好地适应高温、缺水等干旱环境,植物叶片解剖结构均具有相应的变化特征[30-32]。不同干旱条件下的植物,其叶片本身、叶表皮附属结构、机械组织、输导组织及通气组织等都较发达[33]。植物适应干旱环境的第一个特点是尽量缩小叶面积以减少蒸腾量,但不同植物适应干旱环境所采取的策略都有所不同[1]。有些植物随环境干旱程度的加剧,不仅叶面积减小,有时退化形成鳞片状甚至无叶[1,34,35],有些植物叶片较多肉质或薄叶植物的叶片卷曲[6,36,37];或叶片上下表皮角质层加厚[34,37-42],表皮较多白色绒毛,并且密度增加[6],叶表皮蜡质结构增多[35];或叶肉细胞较小,栅栏组织厚度增加,栅海组织比增大以及后壁组织发达[35];或叶脉致密,中脉厚度增加,叶脉维管束及导管腔发达[43];或气孔下陷,单位面积气孔数增多[33]等。而以上适应策略在同种植物中并不是单独存在的,有些植物在极度干旱环境下同时采取以上少数几种或多种适应策略[12]。黄振英等[36]把高温、干旱环境下不同适应策略的植物归纳为薄叶植物、多浆植物、肉茎植物和卷叶植物4类,其叶片可归为正常型、全栅型、环栅型、不规则型、禾草型、退化型6类。

图6 结构相似性系数与遗传相似性系数之间的Mantal检验结果Fig.6 Result of Mantel test between structure indices and genetic indices
短花针茅叶片也具有典型的旱生特征,其适应策略为:1)叶片横切面完全卷曲成“V”形或椭圆形,它属于薄叶卷叶植物,以缩小暴露在空气中的叶面积;2)表皮细胞排列整齐密集,减少蒸散量,并且上表皮有致密的表皮毛,可反射阳光,降低叶表面的温度;3)有发达紧密的维管组织,增强了输导水分和有机物的能力,而维管束导管腔也较发达,可增加对水分的输导能力;4)叶片上表皮弯曲的每个凹陷处有数个运动细胞,可根据外部环境的变化调整叶片的卷曲程度;5)厚壁组织排列紧密,可减小水分蒸腾损失;6)角质层较厚,增加水分的扩散阻力,使叶片叶肉细胞更有效的利用水分,发达的角质层也可保护叶肉组织以免被强光灼伤;7)气孔密度大,由于影响光合作用的一个主要因子为CO2的浓度[31],因此大的气孔密度会增加CO2的吸收量,从而提高光合速率。
3.2 热量是引起短花针茅叶片结构差异的主导气候因子,水分居次要地位
在长期的进化过程中,每一种植物存在于一个适于生长和发育的地理区域内,形成了与气候因素(例如温度、降水、光照等)相适应的分布格局[44]。因此气候是在大尺度上决定物种分布的主要因素[45]。通常认为,决定大尺度植被分布格局及植物形态结构差异的因素主要为温度和降水等气候因子[30,37]。而在小尺度上,影响植物形态结构差异主要由微环境导致,如地形条件(海拔)、土壤母质性质、土壤盐分含量、土壤水分条件及其他生物因素[35,46]。植物的分布与气候因子的关系,尤其是水热因子的关系逐渐从定性描述转变为定量描述[47,48]。本项关于短花针茅叶片解剖结构与气候因子关系的研究与之有相同的结果。在较为广阔的研究区域内(即大的空间尺度),热量因子的分异对短花针茅叶片结构的变化产生了决定性的作用,水分因子处于次要地位,尽管其作用也是不能够被忽视的。
既然热量和水分因子是引起短花针茅叶片解剖结构分化的主要气候因子,那么必定有一种或多种叶片结构与热量和水分因子之间呈现相关关系。研究结果显示,短花针茅叶片维管束/叶面积与热量因子(年平均气温)和水分因子(年降水量)均显著正相关,主脉导管腔/主维管束面积与热量因子(年平均气温)显著正相关,因此认为短花针茅极有可能在适应生长环境的水热变化时,首先采取的策略是改变维管束结构,使得在高温环境下能够吸收更多的水分来调节叶片自身表面的温度[44,49]。
样地8虽然与样地7的地理位置非常相近,但是由于样地8的年平均降水量在8个样地中处于最低水平。因此,样地8的短花针茅叶片结构分化出能够适应该地区极度干旱环境的独特的叶片结构特征。
3.3 叶片解剖结构和分子结构之间有差异
每个生物都有其独特的遗传特性和形态特征,其形态特征是生物遗传特性的外部表象,而遗传多样性决定了外部形态结构千姿百态的特征,因此形态结构的多样化必然会受到遗传分化的制约[50-53]。
通过与张庆等[25]关于短花针茅遗传多样性研究结果的比较发现,8个样地短花针茅的叶片解剖结构与遗传特征之间存在相似性,但不显著。其原因极有可能是遗传特征受到地理位置隔离的影响,受遗传因子基因流的影响比较大[54],而属于外部形态特征的叶片结构,不仅受到大尺度水热条件的影响,还要受到微环境的影响;另一方面,形态特征是遗传特征的外部表象,叶片只是植物器官的一部分,因此植物的遗传特征不只表现在植物叶片上,而是综合表现在根、茎、叶和其他繁殖器官上[55]。
[1] 周云龙.植物生物学[M].北京:高等教育出版社,1999.
[2] 李长复.叶子表面的适应性能[J].生物学通报,1985,(9):3-5.
[3] 李芳兰,包维楷.植物叶片形态解剖结构对环境变化的响应与适应[J].植物学通报,2005,22(B08):118-127.
[4] 冯金朝,周宜君,周海燕,等.沙冬青对土壤水分变化的生理响应[J].中国沙漠,2001,21(3):9-12.
[5] 周宜君,刘春兰,贾晓红,等.沙冬青抗旱、抗寒机理的研究进展[J].中国沙漠,2001,21(3):98-102.
[6] 杨武,郭水良,方芳.不同生境下十七种藓类植物叶的比较解剖学[J].云南植物研究,2007,29(5):409-417.
[7] Parida A K,Das A B,Mittra B.Effects of salt on growth,ion accumulation,photosynthesis and leaf anatomy of the mangrove,B.parviflora[J].Trees-Structure and Function,2004,18(2):167-174.
[8] 陈庆诚,赵松岭,杨凤翔.针茅草原放牧演替中种群消长的数学模型[J].植物学报(英文版),1981,23(4):323-328.
[9] 王勋陵,王静.植物形态结构与环境[M].兰州:兰州大学出版社,1989:1-90.
[10] 周玲玲,宋晓丽.新疆四种补血草属植物叶片的解剖学研究[J].广西植物,2007,27(4):537-559.
[11] 蒋志荣.沙冬青抗旱机理的探讨[J].中国沙漠,2000,20(1):72-75.
[12] Gates D M.Energy,plants,and ecology[J].Ecology,1965,46:1-13.
[13] Gates D M.Energy exchange between organisms and environment[J].Australian Journal of Science,1968,31:67-74.
[14] Gates D M.Transpiration and leaf temperature[J].Annual Review of Plant Physiology,1968,19(1):211-238.
[15] Parkhurst D F,Loucks O L.Optimal leaf size in relation to environment[J].The Journal of Ecology,1972,60:505-537.
[16] Smith W K.Temperatures of desert plants:another perspective on the adaptability of leaf size[J].Science,1978,201:614.
[17] Hegazy A,Elamry M.Leaf temperature of desert sand dune plants:perspectives on the adaptability of leaf morphology[J].African Journal of Ecology,1998,36(1):34-43.
[18] Cunningham S A,Summerhayes B,Westoby M.Evolutionary divergences in leaf structure and chemistry,comparing rainfall and soil nutrient gradients[J].Ecological Monographs,1999,69(4):569-588.
[19] Grubb P J.Plant populations and vegetation in relation to habitat,disturbance and competition:problem of generalization[J].Ecology,1985,63:921-931.
[20] Turner I.Sclerophylly:primarily protective[J].Functional Ecology,1994,8(6):669-675.
[21] 内蒙古植物志编辑委员会.内蒙古植物志(第二版,第五卷)[M].呼和浩特:内蒙古人民出版社,1994:200-202.
[22] 中国科学院内蒙古宁夏综合考察队.内蒙古植被[M].北京:科学出版社,1985.
[23] 卢生莲,吴珍兰.中国针茅属植物的地理分布[J].植物分类学报,1996,34(3):242-253.
[24] 陈世鐄,李银鹏,孟君,等.内蒙古几种针茅特性和生态地理分布的研究[J].内蒙古农牧学院学报,1997,18(1):40-46.
[25] 张庆,牛建明,董建军.内蒙古地区短花针茅(S.breviflora)种群遗传多样性[J].生态学报,2008,28(7):3447-3455.
[26] 李正理.植物制片技术[M].北京:科学出版社,1987.
[27] 李和平.植物显微技术[M].北京:科学出版社,2009.
[28] 康萨如拉,牛建明,张庆.一种植物叶片的解剖结构显微测量新方法——基于GIS技术[J].生物学杂志,2012,29(1):95-96,94.
[29] 牛建明.基于气候的植被空间分布的数字模拟——以内蒙古为例[J].生态学报,2001,21(7):1064-1071.
[30] 胡建莹,郭柯,董鸣.高寒草原优势种叶片结构变化与生态因子的关系[J].植物生态学报,2008,32(2):370-378.
[31] 李翠,程明,唐宇丹,等.青藏高原2种柳属植物叶片解剖结构和光合特征的比较[J].西北植物学报,2009,29(2):275-282.
[32] 孟林,杨宏新,毛培春,等.偃麦草属植物种间苗期抗旱性评价[J].草业学报,2011,20(5):34-41.
[33] 强科斌,郭敏,张晓庆,等.圆囊苔草叶片的形态解剖学研究[J].草业学报,2007,16(2):76-83.
[34] 布仁图雅.内蒙古典型草原退化演替过程中植物个体小型化解剖学机制研究[D].呼和浩特:内蒙古大学,2007.
[35] 邓彦斌,姜彦成,刘健.新疆10种藜科植物叶片和同化枝的旱生和盐生结构的研究[J].植物生态学报,1998,22(2):69-75.
[36] 黄振英,吴鸿,胡正海.30种新疆沙生植物的结构及其对沙漠环境的适应[J].植物生态学报,1997,21(6):34-43.
[37] 蔡永立,王希华,宋永昌.中国东部亚热带青冈种群叶片的生态解剖[J].生态学报,1999,19(6):844-849.
[38] He J S,Wang X L.Morphological and anatomical features of Quercus section suber and its adaptation to the ecological environment[J].Acta Phytoecologica Sinica,1994,18(3):219-227.
[39] 孙宪芝,郑成淑,王秀峰.木本植物抗旱机理研究进展[J].西北植物学报,2007,27(3):629-634.
[40] 杨九艳,杨劫,杨明博,等.鄂尔多斯高原锦鸡儿属植物叶表皮特征及生态适应性[J].植物生态学报,2005,29(6):961-967.
[41] 史刚荣,赵金丽,马成仓.淮北相山不同群落中3种禾草叶片的生态解剖[J].草业学报,2007,16(3):62-68.
[42] 万宏伟,杨阳,白世勤,等.羊草草原群落6种植物叶片功能特性对氮素添加的响应[J].植物生态学报,2008,32(3):611-621.
[43] 李亚男,罗丽娟.不同居群狗牙根的叶片解剖结构比较研究[J].草业学报,2010,19(4):149-158.
[44] Cox C B,Moore P D.Biogeography:an Ecological and Evolutionary Aapproach[M].Oxford:Blackwell Publishing,2010.
[45] 余振,孙鹏森,刘世荣.中国东部南北样带主要植被类型物候期的变化[J].植物生态学报,2010,(3):316-329.
[46] 邰建辉,王彦荣,李晓霞,等.不同覆盖物对无芒隐子草建植的影响[J].草业学报,2011,20(3):287-291.
[47] 柯文山,钟章成,杨毅,等.四川大头茶地理分布与环境因子的关系[J].生态学杂志,1999,18(6):24-27.
[48] 莫亿伟,郭振飞,谢江辉.温度胁迫对柱花草叶绿素荧光参数和光合速率的影响[J].草业学报,2011,20(1):96-101.
[49] 李洪山,张晓岚,侯彩霞,等.梭梭适应干旱环境的多样性研究[J].干旱区研究,1995,12(2):15-17.
[50] Helenurm K,Ganders F R.Adaptive radiation and genetic differentiation in Hawaiian Bidens[J].Evolution,1985,39:753-765.
[51] Gottlieb L,Warwick S,Ford V S.Morphological and electrophoretic divergence betweenLayiadiscoideaandL.glandulosa[J].Systematic Botany,1985,10:484-495.
[52] Charlesworth B,Lande R,Slatkin M.A neo-Darwinian commentary on macroevolution[J].Evolution,1982,36(3):474-498.
[53] Land R,Arnold S J.The measurement of selection on correlated characters[J].Evolution,1983,50:1210-1226.
[54] 王中生,安树青,冷欣,等.岛屿植物舟山新木姜子居群遗传多样性的RAPD分析[J].生态学报,2004,24(3):414-422.
[55] 潘瑞炽,李娘辉.植物生理学[M].北京:高等教育出版社,2001.
猜你喜欢
林业资源管理(2022年6期)2023-01-18
现代园艺(2021年23期)2021-12-01
河北农业大学学报(2021年5期)2021-11-10
林业勘查设计(2020年1期)2021-01-18
新农业(2020年18期)2021-01-07
新疆农业科学(2020年9期)2020-10-13
河南农业科学(2019年9期)2019-09-24
江苏农业科学(2017年5期)2017-04-15
西南农业学报(2016年6期)2016-04-16
儿童故事画报·发现号趣味百科(2015年7期)2015-10-23
