
人高密度脂蛋白受体基因3'UTR 对其mRNA 稳定性的影响
2013-04-20 07:48王丽贾晓健姜华军杜郁杨帆司书毅洪斌
中国医药生物技术 2013年1期
王丽,贾晓健,姜华军,杜郁,杨帆,司书毅,洪斌
心血管疾病是当今世界发病率最高的疾病之一,动脉粥样硬化(atherosclerosis,AS)是心血管疾病的主要病理基础,而脂质代谢紊乱是公认的危险因子。长期的临床研究表明,高密度脂蛋白胆固醇(HDL-C)的降低(< 40 mg/dl)是心血管疾病独立而重要的危险因子。美国心血管疾病药物专家 Rader 提出,降低心血管疾病危害的下一个目标就是寻找能够提高 HDL-C 水平和(或)增强 HDL 功能的药物和治疗方案[1]。研究表明,HDL 最重要的抗动脉粥样硬化机制是促进胆固醇的逆转运(reverse cholesterol transfer,RCT),即胆固醇从外周组织向肝脏转运并排出的过程[2]。高密度脂蛋白受体 SR-BI(人 SR-BI 又称 CLA-1)作为 HDL 受体,通过上调其表达可促进 HDL-C 直接转运至肝脏并最终被排出,加速胆固醇的逆转运,因此,SR-BI 被认为是一个潜在的预防和(或)逆转动脉粥样硬化的新靶点[3-4]。除转录水平的调控外,mRNA 的稳定性相关的转录后水平调控是基因表达水平的另一个重要调控机制,RNA 结合蛋白通过识别特定的 mRNA 3' 非翻译区(3' untranslated region,3'UTR)的顺式作用元件来影响 mRNA 的半衰期,参与 mRNA 稳定性、基因表达定位以及翻译效率等调控[5]。目前,对于基因 mRNA 稳定性的调控已成为药物发现的潜在靶点[6]。
本研究以人 HDL 受体 SR-BI/CLA-1 基因3'UTR 介导的 mRNA 稳定性为靶点,将 SR-BI/ CLA-1 基因 3'UTR 大约 1200 bp 的全长片段克隆至荧光素酶报告基因的下游,构建受 SR-BI/ CLA-1 基因 3'UTR 调控的重组荧光素酶报告质粒 pc-luc-3'UTR 以及 pGL3-CLAp-luc-3'UTR,并获得了稳定转染 pGL3-CLAp-luc-3'UTR 的 HepG2 细胞的单克隆细胞株,为进一步研究 SR-BI/CLA-1 表达调控机制以及针对人 SR-BI/CLA-1 基因 mRNA 稳定性的表达上调剂高通量筛选模型的建立奠定了基础。
1 材料与方法
1.1 材料
1.1.1 质粒、菌株和细胞株 质粒载体 pGEM-T、真核细胞表达载体 pGL3-Basic 和 pcDNA3.1 均 为美国 Promega 公司产品;大肠杆菌 E. coli DH5α 和人肝癌细胞株 HepG2 为本实验室保存。
1.1.2 工具酶和主要试剂 Pfu DNA 聚合酶购自上海生工生物工程公司;T4 DNA 连接酶、限制 性内切酶等均购自大连宝生物工程公司;DNA 回收试剂盒购自北京全式金生物技术有限公司;PureYield™ 中量质粒提取试剂盒、细胞裂解液及荧光素酶检测系统、SV 总 RNA 提取试剂盒均购自美国 Promega 公司;转染试剂 Lipofectamine™ 2000、SuperScript III First-Strand 逆转录试剂盒均购自美国 Invitrogen 公司;实时荧光定量 RT-PCR 试剂盒(FastStart Universal SYBR Green PCR Master)购自瑞士 Roche 公司;MEM 细胞培养基购自美国 Thermo 公司;胎牛血清、丙酮酸钠、非必需氨基酸和 G418 抗生素购自美国 Gibco 公司;二甲基亚砜(DMSO)购自美国 Sigma 公司。 1.1.3 仪器 MiniCycle PTC-200 型 PCR 仪为美国 MJ Research 公司产品;EnVision 多功能微 孔板检测仪为美国 PerkinElmer 公司产品;iQ5 Multicolor real time PCR 仪为美国 Bio-Rad 公司产品。
1.2 方法
1.2.1 CLA-1 基因 mRNA 3'UTR 目的片段的获得 以人肝癌细胞株 HepG2 基因组 DNA 为模板,PCR 扩增获得 CLA-1 基因 mRNA 3'UTR 全长序列(1 ~ 959 bp,1 为终止密码子),上、下游引物分别为 5' GCTCTAGATGCCTTTCTCCACAG GGTCC 3',5' GCTCTAGA GGGCGTACTCATGCAG TAT 3'(下划线为 Xba I 酶切位点)。PCR 体系反应条件为 94 ℃ 变性 5 min;94 ℃ 变性 50 s, 60 ℃ 退火 50 s,72 ℃ 延伸 60 s(30 个循环);72 ℃ 10 min。将 PCR 扩增产物回收纯化并连接至 pGEM-T 载体,转化 E. coli DH5α 感受态宿主菌,通过蓝白斑筛选获得重组单克隆。提取纯化质粒,酶切鉴定阳性克隆,将该重组质粒命名为 pGEM-3'UTR,测序鉴定。
1.2.2 重组质粒的构建和鉴定 将 pGL3-Basic 质粒通过 Hind III 和 Xba I 双酶切获得荧光素酶报告基因的 DNA 序列,并连接至 pcDNA3.1 载体多克隆位点,获得重组质粒 pc-luc。将上述获得的测序正确的 CLA-1 mRNA 3'UTR 片段克隆至 pc-luc 质粒中荧光素酶报告基因的下游 Xba I 位点处,经插入方向鉴定正确后最终获得重组质粒 pc-luc-3'UTR。与此同时,将该 CLA-1 mRNA 3'UTR 片段克隆至之前构建的重组质粒 pGL3-CLAp-luc[7]中荧光素酶报告基因的下游 Xba I 位点处,获得同时克隆有 CLA-1 基因启动子区(CLAp)和其 3'UTR 片段的重组荧光素酶报告质粒 pGL3- CLAp-luc-3'UTR,转染后获得稳定转染细胞株 CLAp-luc-3'UTR HepG2。该质粒同时可检测化合物对 CLA-1 启动子活性或 3'UTR 相关的 mRNA 稳定性的影响,因此后续模型的建立在该质粒的基础上进行。
1.2.3 细胞培养及转染 HepG2 细胞培养于含 10% 胎牛血清、1 mmol/L 丙酮酸钠和 1% 非必需氨基酸的 MEM 培养基中,于 37 ℃、5% CO2条件下培养。0.25% 胰酶加 0.02% 乙二胺四乙酸(EDTA)消化传代。转染用质粒的提取方法按 PureYield™ 中量质粒提取试剂盒说明书进行。将 1 × 105个HepG2 细胞接种于 30 mm 培养皿中,待细胞贴壁生长汇聚程度达到 80% 左右时,利用脂质体 Lipofectamine™2000 将 pc-luc-3'UTR 瞬 时转染至 HepG2 细胞,转染 6 h 后更换培养液,24 h 后对荧光素酶活性进行检测,以确定最优转染效率。
1.2.4 稳定转染细胞株的构建 将瞬时转染 pGL3-CLAp-luc-3'UTR 质粒的细胞以 5 × 103个细胞/孔接种于 96 孔板,于 37 ℃、5% CO2条件下进行培养,以含 700 μg/ml G418 的 MEM 选择培养液筛选稳定转染细胞株,每 3 天更换培养液。 2 周后,选择存活的细胞集落进行扩大培养,并测定荧光素酶的活性,将活性较高的细胞集落以 1∶2 的比例进行分级稀释,筛选稳定转染的单克隆细胞株,传代并进行观察和荧光素酶活性测定,最终获得呈现正常细胞周期、能够稳定传代 20 代以上的高表达荧光素酶活性的细胞株,命名为 CLAp-luc- 3'UTR HepG2。
1.2.5 实时荧光定量 PCR 待测化合物作用于 HepG2 细胞后,提取细胞的总RNA,采用试剂盒 SuperScript III First-Strand 合成 cDNA,采用 2 × TaqMan®gene expression master mix 进行荧光定量 PCR 反应,利用软件分析并计算 Ct 值。以 GAPDH 为内参,采用相对 Ct 的方法对荧光素酶基因的转录水平进行定量。
1.2.6 基因 mRNA 半衰期测定 待测化合物作用细胞 10 h 后在培养基中分别加入 5 μg/ml 放线菌素 D 以终止转录。加入放线菌素 D 后的 0、30、60、90 min 分别提取细胞总 RNA,进行定量 分析。待测基因 mRNA 水平与时间的相关性,其斜率为降解常数 kd,由此计算待测基因 mRNA 的半衰期(t1/2= 0.693/kd)。
2 结果
2.1 CLA-1 基因 mRNA 3'UTR 序列的获得
由于 CLA-1 基因 3'UTR 位于一个外显子中,因此我们以人基因组 DNA 为模板,PCR 扩增得到 CLA-1 基因 mRNA 3'UTR 序列,扩增产物经琼脂糖凝胶电泳检测,显示获得预期大小的 DNA 片段(图 1A)。将该 PCR 产物克隆至 pGEM-T 载体中,并对该插入片段进行序列测定,结果表明 该片段与 GenBank 中报道的序列一致,符合预期设计。
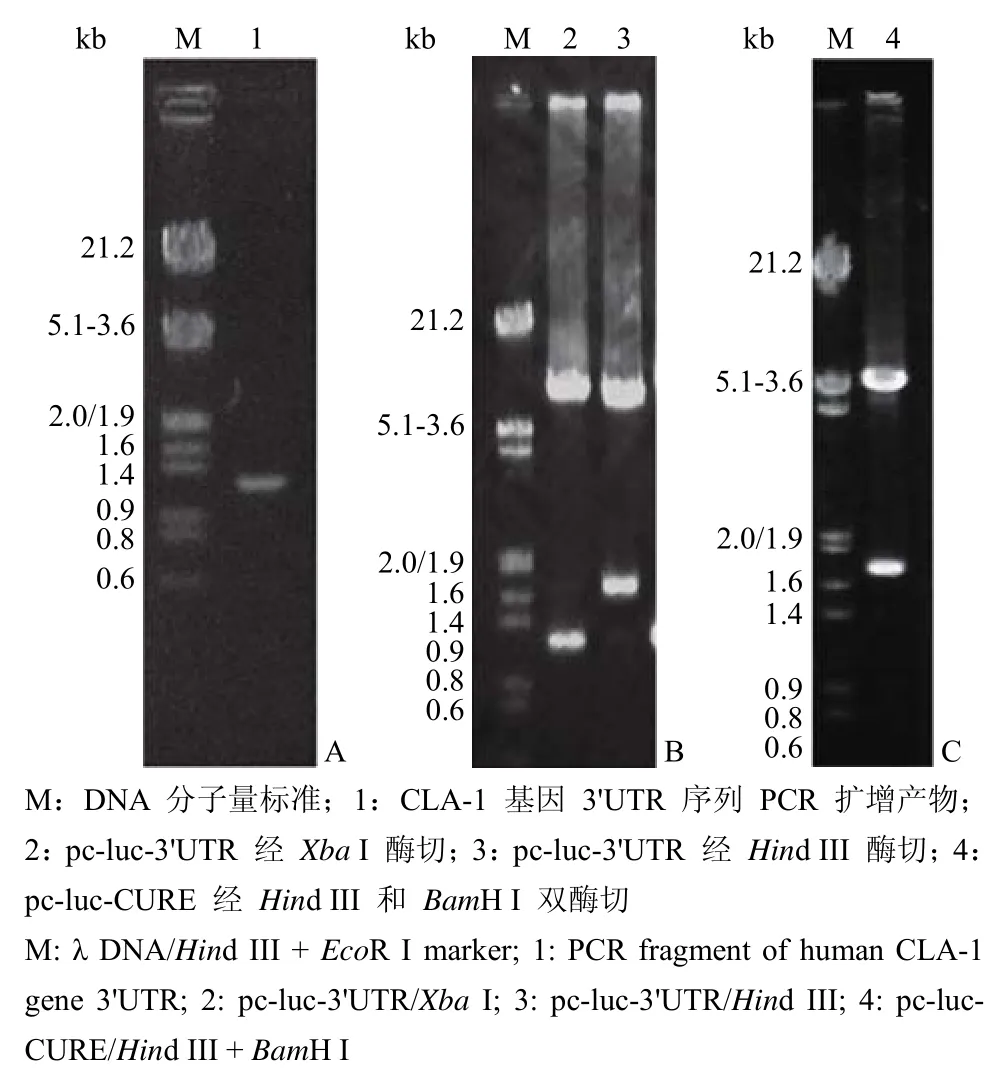
图1 CLA-1 基因 3'UTR 序列的 PCR 扩增产物(A)和 pc-luc-3'UTR(B)、pc-luc-CURE(C)酶切鉴定结果 Figure 1 PCR product of human CLA-1 gene mRNA 3'UTR (A) and restriction enzymatic analysis of recombinant plasmid pc-luc-3'UTR (B) and pc-luc-CURE (C)
2.2 报告基因重组质粒 pc-luc-3'UTR 的构建
对构建的重组质粒 pc-luc-3'UTR 进行酶切鉴定,结果显示 CLA-1 基因 mRNA 3'UTR 序列以正确的方向插入到 pc-luc 中荧光素酶报告基因的下游 Xba I 位点处(图 1B),构建示意图见图 2A。其中,不含 3'UTR 的质粒 pc-luc 作为对照质粒。另外,为了验证 CLA-1 基因 mRNA 3'UTR 的作 用,根据文献[8]报道人工合成了一段已知的保守的 CURE(CU-rich element)序列,构建了 3' 端含有 CURE 的荧光素酶报告质粒 pc-luc-CURE,酶切鉴定结果如图 1C 所示。上述重组质粒均以 pcDNA3.1 质粒自身强启动子 pCMV 为启动子。
2.3 CLA-1 基因 3'UTR 对 mRNA 稳定性的影响
将含有 CLA-1 基因 mRNA 3'UTR 序列的重组质粒 pc-luc-3'UTR 转染HepG2 细胞后检测荧光素酶活性,结果显示,与对照质粒 pc-luc 相比,pc-luc-3'UTR 转染细胞的荧光素酶活性显著降低,而 pc-luc-CURE 则未见明显变化(图 2B)。此外,我们利用实时荧光定量 RT-PCR 的方法检测了分别转染pc-luc、pc-luc-3'UTR 和 pc-luc-CURE 后 HepG2 细胞的荧光素酶 mRNA 水平,结果与荧光素酶活性结果一致(图 2C)。进而对 pc-luc-3'UTR 转染 HepG2 细胞的荧光素酶的半衰期进行了检测,结果显示 CLA-1 3'UTR 的存在显著降低了荧光素酶 mRNA 的稳定性,半衰期由 81.5 min 缩短至 44.4 min(图 2D)。综上结果表明,CLA-1 基因 3'UTR 对荧光素酶活性及其 mRNA 水平和稳定性有显著影响。
2.4 稳定转染细胞株 CLAp-luc-3'UTR HepG2 的建立和应用
由于自身启动子可更好地反映细胞中基因的表达调控状况,于是我们将CLA-1 基因 3'UTR 克隆至先前构建的带有 CLA-1 启动子的荧光素酶报告质粒 pGL3-CLAp-luc[6]中荧光素酶报告基因下游,构建重组报告质粒 pGL3-CLAp-luc-3'UTR,并建立稳定转染 pGL3-CLAp-luc-3'UTR 的 HepG2 细胞,以 CLAp-luc HepG2 细胞作为对照。重组质粒构建示意图见图 3A、3B。
进而采用荧光素酶报告基因和实时荧光定量 RT-PCR 的方法对构建的 CLAp-luc-3'UTR 细胞和 CLAp-luc 对照细胞的荧光素酶活性、报告基因 mRNA 水平及半衰期进行了检测,结果如图 3 所示。CLA-1 基因 3'UTR 的存在可显著降低模型荧光素酶活性及其 mRNA 水平,同时荧光素酶报告基因的半衰期也从 16.2 h 缩短至 2.2 h,与前期构建的 pc-luc-3'UTR 模型结果基本一致。上述结果确证了 CLA-1 基因 3'UTR 的序列中存在对其 mRNA 稳定性有显著影响的顺式元件,使其 mRNA的降解率大大提高。综上结果,本文证实了 CLA-1 基因 3'UTR 的存在对其 mRNA 稳定性的影响。
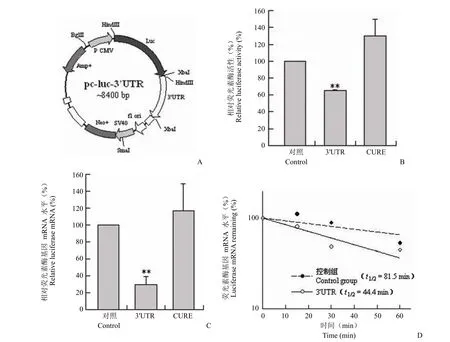
图2 CLA-1 基因 3'UTR 对 mRNA 稳定性的影响(A:重组质粒 pc-luc-3'UTR 构建示意图;B:CLA-1 基因 mRNA 3'UTR 降低荧光素酶的活性;C:CLA-1 基因 mRNA 3'UTR 降低荧光素酶基因 mRNA 的水平;D:CLA-1 基因 mRNA 3'UTR 降低荧光素酶基因 mRNA 的稳定性) Figure 2 The effect of CLA-1 gene 3'UTR on luciferase mRNA stability (A: Illustration of recombinant plasmid pc-luc-3'UTR construction; B: The effect of CLA-1 gene 3'UTR on luciferase activity; C: The effect of CLA-1 gene 3'UTR on luciferase mRNA level; D: The effect of CLA-1 gene 3'UTR on luciferase mRNA stability)
3 讨论
心血管疾病是全球范围造成死亡的首要原因,动脉粥样硬化以富含脂肪的斑块在大动脉壁聚积为主要特征。尽管他汀类药物能减少约 30% 的冠状动脉事件[9],但每年仍有 9% 的患者受到残余心血管病变风险的困扰[10],因此亟需新作用机制调血脂药物的研发。近年来,SR-BI 的抗动脉粥样硬化作用成为研究热点,相关研究逐步揭示肝上 HDL 受体介导的 HDL-C 的选择性摄取将外周组织中的胆固醇转运到肝脏进行处置代谢,是胆固醇逆转运机制的终末步骤,也是最重要的步骤之一,通过上调其表达可促进 HDL-C 直接转运至肝脏并最终被排出,加速胆固醇的逆转运。因此,SR-BI 被认为是一个潜在的预防和(或)逆转动脉粥样硬化的新的治疗靶点[1,11]。
SR-BI 在包括肝脏在内的多种组织中表达,在巨噬细胞上也有表达,但在细胞胆固醇外排中的作用尚存在争议。SR-BI 在肝脏中的基因表达调控在脂代谢过程中尤为重要,受到饮食、激素、胆固醇代谢及药物等的调节。目前针对 SR-BI 基因表达调控机制的研究主要集中在基因转录水平的调控,其中 SREBP、Sp1、LRH-1 等转录因子和 PPARγ、PXR、FXR、LXR 等核受体在调节 SR-BI 表达的过程中发挥了重要的作用[12]。基于此,我们在之前工作中构建了 SR-BI/CLA-1 转录上调剂筛选模型,经筛选获得了以 TSA 为代表的多个活性化合物,并进一步在细胞水平上对其 CLA-1 表达上调活性及其作用机制进行了研究[13-14]。针对 SR-BI/ CLA-1 基因转录后水平的研究尚未见报道。

图3 CLA-1 基因 3'UTR 对 mRNA 稳定性的影响(A:对照质粒 pGL3-CLAp-luc 构建示意图;B:重组质粒 pGL3-CLAp-luc-3'UTR 构建示意图;C:CLA-1 基因 mRNA 3'UTR 降低荧光素酶的活性;D:CLA-1 基因 mRNA 3'UTR 降低荧光素酶基因 mRNA 的水平;E:CLA-1 基因 mRNA 3'UTR 降低荧光素酶基因 mRNA 的稳定性) Figure 3 The effect of CLA-1 gene 3'UTR on luciferase mRNA stability (A: Illustration of control plasmid pc-luc construction; B: Illustration of recombinant plasmid pGL3-CLAp-luc-3'UTR construction; C: The effect of CLA-1 gene 3'UTR on luciferase activity; D: The effect of CLA-1 gene 3'UTR on luciferase mRNA level; E: The effect of CLA-1 gene 3'UTR on luciferase mRNA stability)
与基因转录调控相比,目前人们对于转录后调控细节还所知甚少,但这方面的研究正日益受到重视。人 LDL 受体(LDLR)的 3'UTR 对其 mRNA 稳定性起重要作用,研究发现天然产物小檗碱能够通过作用于人 LDLR 的 3'UTR 从而增加 LDLR 基因 mRNA 的稳定性,以不同于他汀类药物的作用机制提高 LDLR 的表达水平,降低血浆中 LDL-C 水平,有可能发展为新型降血脂药物,并可与他汀类药物联用[6]。另外,Benjamin 等[15-16]利用绿色荧光蛋白作为报告基因构建了克隆有 IL-3、TNFα 基因 3'UTR 的重组报告质粒,以此为基础获得可增加这些细胞因子 mRNA 稳定性的表达调控剂。已报道的转录后调控相关的顺式作用元件包括 ARE(AU-rich element)、CURE、GRE(GU-rich element)、PRE(pyrimidine-rich element)等。RNA 结合蛋白通过识别特定的 3'UTR 顺式作用元件来决定 mRNA 的半衰期和降解率,参与 mRNA 的稳定性、基因表达的定位以及翻译效率等生理过程的调控[5]。此外,近年来研究发现 microRNA(miRNA)可通过与靶基因 mRNA 的 3'UTR 上的特定区域通过完全或不完全碱基互补结合,介导 mRNA 的降解或翻译抑制,从而在转录后水平调控靶基因的表达,成为转录后水平调控研究的新热点。最新报道发现 miR-33 可通过直接靶向 ABCA1/G1 基因 3'UTR 区,在转录后水平抑制 ABCA1 和 ABCG1 基因的表达,从而调控了细胞内胆固醇代谢平衡[17-19]。因此,转录后调控是除转录水平调控外的另一个重要的基因表达调控机制,已成为药物发现的重要靶点。
人 HDL 受体基因 CLA-1 转录并加工成熟的 mRNA 全长 2759 bp,其3'UTR 序列长 1234 bp,由第 13 个外显子编码。不同种属的 SR-BI 基因 3'UTR 区域中存在高度保守序列,提示其具有转录后调节机制。本研究结果显示,将 CLA-1 基因 3'UTR 构建至荧光素酶报告基因 3' 端,可明显降低荧光素酶活性及基因 mRNA 的稳定性(图 3),说明除转录水平的调控外,SR-BI/CLA-1 基因 mRNA 3'UTR 介导的 mRNA 的稳定性相关的转录后调控也是其基因表达水平的另一个重要调控机制,可以作为上调 CLA-1 表达的一个新靶点,为后续针对 SR-BI mRNA 稳定性为靶点的模型建立提供了依据。
以本研究获得的稳定转染细胞 CLAp-luc- 3'UTR HepG2 为基础建立筛选模型,不仅可以获得上调 CLA-1 启动子活性的化合物,同时还可获得增加 CLA-1 基因 3'UTR 介导的 mRNA 的稳定性的化合物,这为 CLA-1 表达上调剂的发现提供了更为有效可靠的基础。另外也可用于参与调控 SR-BI/CLA-1 表达的 miRNAs 筛选和作用机制研究。这些活性化合物或 miRNAs 的发现,一方面可以大大丰富 CLA-1 基因表达上调剂的候选物,并有可能和 CLA-1 基因转录上调剂联合作用,更好地上调 CLA-1 基因的表达,为最终发现高效、低毒的基于 HDL 的抗动脉粥样硬化药物奠定基础;另一方面,这些活性化合物或 miRNAs 还将有助于深入探讨 CLA-1/SR-BI 基因 3'UTR 的转录后调控机制,对于探索开发新型基于 HDL 抗动脉粥样硬化药物具有重要的科学意义和应用价值。
[1] Rader DJ. Molecular regulation of HDL metabolism and function: implications for novel therapies. J Clin Invest, 2006, 116(12):3090- 3100.
[2] Fredenrich A, Bayer P. Reverse cholesterol transport, high density lipoproteins and HDL cholesterol: recent data. Diabetes Metab, 2003, 29(3):201-205.
[3] Acton S, Rigotti A, Landschulz KT, et al. Identification of scavenger receptor SR-BI as a high density lipoprotein receptor. Science, 1996, 271(5248):518-520.
[4] Calvo D, Vega MA. Identification, primary structure, and distribution of CLA-1, a novel member of the CD36/LIMPII gene family. J Biol Chem, 1993, 268(25):18929-18935.
[5] Hughes TA. Regulation of gene expression by alternative untranslated regions. Trends Genet, 2006, 22(3):119-122.
[6] Kong W, Wei J, Abidi P, et al. Berberine is a novel cholesterol- lowering drug working through a unique mechanism distinct from statins. Nat Med, 2004, 10(12):1344-1351.
[7] Yang Y, Zhang Z, Jiang W, et al. Identification of novel human high-density lipoprotein receptor Up-regulators using a cell-based high-throughput screening assay. J Biomol Screen, 2007, 12(2):211- 219.
[8] Wang S, Zhang J, Theel S, et al. Nitric oxide activation of Erk1/2 regulates the stability and translation of mRNA transcripts containing CU-rich elements. Nucleic Acids Res, 2006, 34(10):3044-3056.
[9] Brugts JJ, Yetgin T, Hoeks SE, et al. The benefits of statins in people without established cardiovascular disease but with cardiovascular risk factors: meta-analysis of randomised controlled trials. BMJ, 2009, 338:b2376.
[10] Hausenloy DJ, Yellon DM. Targeting residual cardiovascular risk: raising high-density lipoprotein cholesterol levels. Heart, 2008, 94(6):706-714.
[11] Acton SL, Kozarsky KF, Rigotti A. The HDL receptor SR-BI: a new therapeutic target for atherosclerosis? Mol Med Today, 1999, 5(12): 518-524.
[12] Yang Y, Wang L, Si S, et al. How can high-throughput screening deliver drugs to treat atherosclerosis? Expert Opin Drug Discov, 2010, 5(12):1175-1188.
[13] Yang Y, Jiang W, Wang L, et al. Characterization of the isoflavone pratensein as a novel transcriptional up-regulator of scavenger receptor class B type I in HepG2 cells. Biol Pharm Bull, 2009, 32(7):1289-1294.
[14] Bao Y, Yang Y, Wang L, et al. Identification of trichostatin A as a novel transcriptional up-regulator of scavenger receptor BI both in HepG2 and RAW 264.7 cells. Atherosclerosis, 2009, 204(1):127-135.
[15] Benjamin D, Colombi M, Moroni C. A GFP-based assay for rapid screening of compounds affecting ARE-dependent mRNA turnover. Nucleic Acids Res, 2004, 32(11):e89.
[16] Benjamin D, Colombi M, Stoecklin G, et al. A GFP-based assay for monitoring post-transcriptional regulation of ARE-mRNA turnover. Mol Biosyst, 2006, 2(11):561-567.
[17] Rayner KJ, Suárez Y, Dávalos A, et al. MiR-33 contributes to the regulation of cholesterol homeostasis. Science, 2010, 328(5985):1570- 1573.
[18] Najafi-Shoushtari SH, Kristo F, Li Y, et al. MicroRNA-33 and the SREBP host genes cooperate to control cholesterol homeostasis. Science, 2010, 328(5985):1566-1569.
[19] Marquart TJ, Allen RM, Ory DS, et al. miR-33 links SREBP-2 induction to repression of sterol transporters. Proc Natl Acad Sci U S A, 2010, 107(27):12228-12232.
猜你喜欢
天津医科大学学报(2021年4期)2021-08-21
昆明医科大学学报(2021年8期)2021-08-13
昆明医科大学学报(2021年3期)2021-07-22
西安交通大学学报(医学版)(2021年3期)2021-05-15
中日友好医院学报(2021年1期)2021-04-14
广东蚕业(2021年1期)2021-03-18
临床超声医学杂志(2020年12期)2020-12-20
中西医结合肝病杂志(2020年2期)2020-10-27
山东医药(2020年9期)2020-05-20
热带农业科学(2015年9期)2015-10-14
