
云南汉族原发性肌张力障碍患者TOR1A基因突变研究☆
2013-09-17 08:31隆昱洲雷进罗丽华陈婷婷孙倩倩褚嘉祐杨昭庆孙浩
中国神经精神疾病杂志 2013年9期
隆昱洲雷进罗丽华陈婷婷孙倩倩褚嘉祐杨昭庆孙浩
肌张力障碍(dystonia)是一种不自主、持续性肌肉收缩引起的扭曲、重复运动或姿势异常的综合症[1]。临床上,症状单一(震颤除外)并无神经退化或其它获得性因素引起的肌张力障碍,一般称为原发性肌张力障碍(primary dystonia)[2]。或者根据发病年龄可分为早发型(26岁前发病)和迟发型(26岁后发病)[1,3]。随着对引起肌张力障碍发生遗传因素的不断研究,已经发现20多个基因座位与其发生相关[4-5]。其中最早发现的TOR1A(也称为DYT1)基因在早发型肌张力障碍中占有非常重要的作用[6],其第五外显子区的GAG三核苷酸删除(904_906delGAG,GenBank序号:rs80358233)是引起早发型的肌张力障碍的主要遗传因素[7]。但现在对在迟发型的肌张力障碍患者中此基因的突变及其是否有群体特异性情况仍不清楚。因此本研究对122例原发性肌张力障碍患者及100名正常对照TOR1A的第五外显子进行测序,了解此基因突变在中国西南地区汉族肌张力障碍患者中的分布情况,并寻找此基因第五外显子有无新的可能致病突变。
1 对象与方法
1.1 研究对象 患者组为2011年至2012年来我院进行检查的肌张力障碍患者。纳入标准:①符合Geyer等[1]2006年建议诊断标准及原发性肌张力障碍的定义[2];②症状单一,无神经退化或其他获得性因素引起的原发性肌张力障碍。排除标准:其他症状叠加的肌张力障碍患者;或获得性/外生的(acquired/exogenous)的肌张力障碍患者。共收集原发性肌张力障碍患者122例,年龄13~83岁,平均(53.9±14.9)岁,男42例,女80例。对照组来自本院体检者。纳入标准:年龄>50周岁,无神经退行性病变症状。共收集100名,年龄50~65岁,平均(56.5±4.3)岁,男40名,女60名。两组年龄(t=-1.62,P=0.108)、性别分布(χ2=0.73,P=0.39)无统计学差异。
所有研究对象均对本研究知情同意。本研究经中国医学科学院医学生物学研究所伦理委员会同意。
1.2 研究方法
1.2.1 患者分类 收集患者发病年龄、病程长短及发病部位等信息。并根据一般国际通行的标准[1,8],以26岁为界限,将肌张力障碍患者分为早发型(≤26岁)和晚发型(>26岁)两类。
1.2.2 TOR1A基因第五外显子测序 采集所有研究对象外周血5 mL,并用肝素钠抗凝。各血液样本取250µL,使用AxyPrep血基因组DNA小量制备试剂盒(AxyGen,中国)进行DNA抽提。使用引物F:5'-GGTAGGTAAGTGACCCCCAG-3'和 R:5'-AAAAAACACACACACAAGGC-3'对目的区域进行扩增。扩增条件为(PCR试剂来自TaKaRa公司):DNA 样本 20 ng,MgCl2(25 mmol/L)1.2 µL;dNTP(2.5 mmol/L)1.6 µL;正反向引物(5 µmol/L)各1 µL;TaKaRaTaq酶(55 U/µL)0.16 µL;10×PCR Buffer2.5 µL;加水至总体积20 µL。扩增条件为:94℃15 s,60℃30 s,72℃15 s,重复30个循环,72℃10 min。扩增目的片段包括TOR1A基因(GenBank序号:NM_000113)第五外显子5'端81 bp的内含子区域;覆盖了第五外显子的所有氨基酸编码区,并包含其3'非翻译区420 bp长度区域。扩增片段共长752bp。
扩增PCR产物用Wizard®SV Gel and PCR Clean-Up System PCR纯化试剂盒(Promega,美国)进行纯化。纯化产物用BigDye Terminator v1.1试剂盒(Applied Biosystems,美国),分别用上述PCR引物进行双向测序反应。测序反应产物在3130 DNA测序仪(Applied Biosystems,美国)进行电泳,结果用Sequencer Analysis V5软件(Applied Biosystems,美国)和软件Chromas进行分析。
1.3 统计学方法 获得测序结果后,参考数据库中原序列(NM_000113.2)划定第五外显子范围,并对比患者组与对照组之间的编码蛋白质的核苷酸有无改变。确定在本研究群体中突变情况。并计算其突变频率。所计量资料用“±s”描述,计数资料用构成比描述,并采用SPSS 12.0进行统计学分析。对于在基因3'非翻译区检测到的突变用TargetScan软件[9],根据已知miRNA与基因3'作用的规律,预测突变是否对miRNA的结合位点发生影响。并根据突变所在DNA区段在不同物种中的保守性,评估此突变是否可能是潜在的miRNA结合位点,是否可能影响基因表达情况。
2 结果
2.1 患者特征与比例 122例肌张力障碍患者中,早发型肌张力障碍患者占7.4%(9/122)。在迟发型患者中,最多为节段性颅颈肌张力障碍(Meige综合症),占40.7%(46/113);其次为局部性眼睑痉挛(38.9%,44/113)和局部性痉挛性斜颈(15.0%,17/113)(见表1)。
2.2 TOR1A基因第五外显子测序 对TOR1A基因第五外显子进行双向测序显示,8例患者此区域有突变,共检测到5种突变,其中包括多在早发型肌张力障碍患者中发现的突变904_906delGAG及数据库中未收录的突变m.1378TT>TA。对照组未发现904_906delGAG及m.1378TT>TA突变。患者突变类型与症状,见表2。904_906delGAG突变的纯合子、杂合子以及正常对照序列的反向测序结果见图1 A。新发现未收录入库的突变m.1378TT>TA的双向测序结果列于图1 B中。
2.3 突变频率及miRNA作用位点预测 在9例早发型肌张力障碍患者中,1例携带904_906del-GAG突变;迟发型患者中,携带904_906delGAG突变的患者占2.7%(3/113)。在所有肌张力障碍患者中共有4例携带此突变,另有1例为纯合子,此突变在肌张力障碍患者中的基因频率为2.1%(5/244)。
对位于TOR1A基因3'非翻译区所检测到的突变,根据已知miRNA与基因相互作用规律进行推测,并据突变所在DNA区段在不同物种中的保守性比对,数据库中收录的3个SNP(rs75881350,rs1182和rs1183)均未发现这3个SNP与已知miRNA 结合的可能;新发现的 SNPm.1378TT>TA 在人、黑猩猩、大猩猩、猕猴、鼠和兔等哺乳动物中保守,并且其可能为miRNA hsa-miR-494的潜在结合位点。
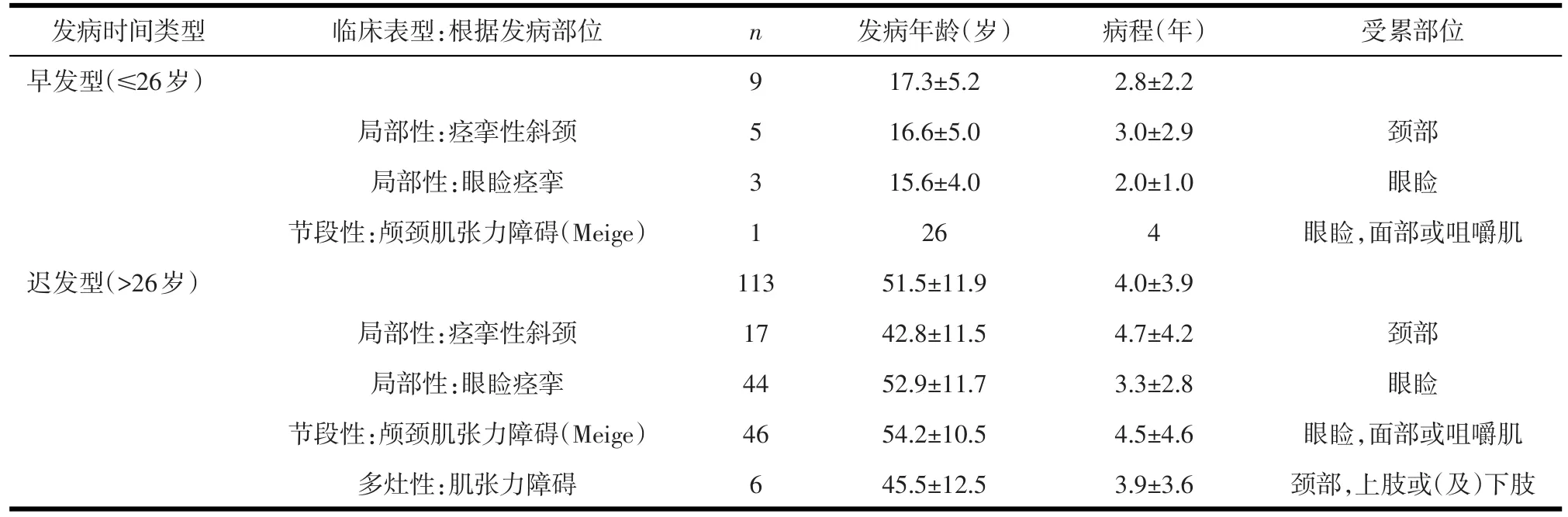
表1 122例肌张力障碍患者分类

表2 8例有突变患者的突变类型及症状
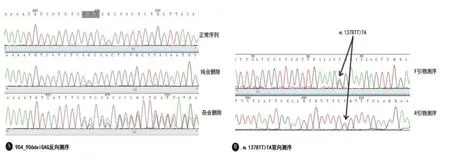
图1 突变测序图 A:为904_906delGAG突变反向测序图,示正常序列、突变纯合子及杂合子,图中黑框为删除的三碱基GAG互补序列CTC;B:为突变m.1378TT>TA双向测序图,箭头所指为突变位点
3 讨论
TOR1A基因(也称DYT1)编码蛋白torsinA,此基因在脑中各部位广泛表达,特别是在中脑灰质区。TOR1A编码蛋白为腺苷三磷酸酶(ATPase)AAA+超家族中的一员[2]。此家族基因参与膜融合,蛋白水解和DNA复制等多种细胞生物学过程,其功能主要是与蛋白的折叠有关,也有学者认为它们是一种分子伴侣[10]。在体外实验中发现,带有904_906delGAG的基因的突变torsinA蛋白(DeltaE302)会导致蛋白形态的改变,使得这一原本位于内质网中的蛋白出现于核膜上[11]。同时这一突变蛋白也使得内质网形态改变。这些改变引起了多巴胺释放神经细胞的损伤,从而可能导致肌张力障碍。在一些小鼠模型中也发现,此基因的过表达也可引起小鼠的运动异常[12]。
尽管体外及体内的实验均指出此基因突变可能为肌张力障碍发病的原因,但是此突变在肌张力障碍发病遗传因素中所占比例,及此基因是否还有其它突变也能引起肌张力障碍仍有待讨论。最初的研究显示[13],904_906delGAG突变在德裔犹太人的早发型肌张力障碍患者中出现的频率非常高(大于90%),因此人们认为其可能是早发型原发性肌张力障碍的主要遗传病因。但随着对多个群体的研究以及对德裔犹太人起源的辨认,人们发现在不同的群体中早发型原发性肌张力障碍患者携带此致病突变的比例有群体特异性;德裔犹太人携带高频的904_906delGAG突变,可能是因为群体遗传学上奠基者效应的影响。我们综合对比这些数据(见表3),可见在欧洲群体中英法德地区[14-16],此突变在早发型患者中所占比例较高,通常大于30%。而在另一些欧洲群体[17-18]及大部分亚洲群体中[19-22],904_906delGAG突变在早发型肌张力障碍患者中所占比例只有10~20%左右。特别是在中国汉族群体中,无论是台湾[22]、吉林[21]还是本研究中对云南汉族的研究均显示,此突变在早发型肌张力障碍患者中所占比例在10%左右。因此,我们认为在中国人群中此突变并不能解释大部分早发型原发性肌张力障碍的病因。中国汉族人群可能携带有其它非此突变的遗传因素而导致早发型原发性肌张力障碍,对于中国人群早发型原发性肌张力障碍的遗传病因仍然值得探讨。

表3 突变类型及携带者症状
讨论了904_906delGAG突变导致早发型原发性肌张力障碍后,世界各地的研究小组也对其在迟发型原发性肌张力障碍中的出现频率进行了讨论。这些结果同样列于表3中。在大部分研究群体中,均没有在迟发型肌张力障碍患者中检测到904_906delGAG突变。只在德国及日本样本中各检测到1例,其频率均小于1%[15,20]。但在本研究中,在113例迟发型肌张力障碍患者中检测到3例携带此突变的患者(2.7%)。此频率高于其它报道人群。虽然可能因样本量小带来偏倚,但本研究仍说明了此突变能导致迟发型肌张力障碍,并且在中国西南迟发型肌张力障碍患者群体中频率略高。这一现象在已报道的人群中较为少见,因此我们推测在云南汉族群体中是否有可能存在一种特殊机制,使肌张力障碍的发病“推迟”。云南地理位置特殊,处于高原低氧及湿热环境,并且云南的汉族人群有特殊的人口历史学来源。这些特殊的环境及遗传因素与904_906delGAG突变致病关系的揭示,可能为肌张力障碍发生的分子机制提供有益帮助。
本研究发现1例纯合型的904_906delGAG突变患者,此患者中突变TOR1A基因的表达量有可能为携带杂合突变患者的两倍。但从症状来看,携带纯合突变病人与其它携带杂合突变的患者差别并不大,如:发病年龄仍为30岁,并没有显著提前。因此我们推测,904_906delGAG突变致肌张力障碍发生的病理机制可能较为复杂,突变基因表达量和疾病的关系有待进一步研究。
本研究还发现位于TOR1A基因3'非翻译区的一个新突变:m.1378 TT>TA。经计算机模拟分析发现,此位点在哺乳动物中保守,并可能是hsamiR-494的潜在结合位点。如其突变可能影响到miR-494的结合,从而引起TOR1A基因转录水平的改变,此突变有可能是一个导致肌张力障碍发生的遗传因素。小鼠实验模型也显示[12],TOR1A的表达量的改变能引起小鼠运动障碍。仅仅根据本研究的病例对照对比,对突变m.1378 TT>TA在引起肌张力障碍中的作用仍不能下结论。但此突变与TOR1A基因的表达关系,及其与肌张力障碍发生的关系在将来也非常值得研究的。
综上所述,904_906delGAG也是导致云南汉族人群中迟发型肌张力障碍发生的遗传因素。并且云南群体可能存在特异的肌张力障碍遗传变异谱。更大规模的肌张力障碍患者遗传学筛查,及更深入的机制研究仍是非常值得进行的。
[1]Geyer HL,Bressman SB.The diagnosis of dystonia[J].Lancet Neurol,2006,5(9):780-790.
[2]Bressman SB.Genetics of dystonia:an overview[J].Parkinsonism Relat Disord,2007,13(S 3):S347-355.
[3]张晓洁,陈晟,乐卫东.肌张力障碍相关基因及临床特征[J].上海交通大学学报(医学版),2008,28(07):908-911.
[4]Spatola M,Wider C.Overview of primary monogenic dystonia[J].Parkinsonism Relat Disord,2012,18(S 1):S158-161.
[5]Petrucci S,Valente EM.Genetic issues in the diagnosis of dystonias[J].Front Neurol,2013,4(1):34.
[6]Ozelius L,Kramer PL,Moskowitz CB,et al.Human gene for torsion dystonia located on chromosome 9q32-q34[J].Neuron,1989,2(5):1427-1434.
[7]Ozelius LJ,Hewett JW,Page CE,et al.The early-onset torsion dystonia gene(DYT1)encodes an ATP-binding protein[J].Nat Genet,1997,17(1):40-48.
[8]de Carvalho Aguiar PM,Ozelius LJ.Classification and genetics of dystonia[J].Lancet Neurol,2002,1(5):316-325.
[9]Garcia DM,Baek D,Shin C,et al.Weak seed-pairing stability and high target-site abundance decrease the proficiency of lsy-6 and other microRNAs[J].Nat Struct Mol Biol,2011,18(10):1139-1146.
[10]Ogura T,Wilkinson AJ.AAA+superfamily ATPases:common structure--diverse function[J].Genes Cells,2001,6(7):575-597.
[11]Goodchild RE,Dauer WT.Mislocalization to the nuclear envelope:an effect of the dystonia-causing torsinA mutation[J].Proc Natl Acad Sci U S A,2004,101(3):847-852.
[12]Shashidharan P,Sandu D,Potla U,et al.Transgenic mouse model of early-onset DYT1 dystonia[J].Hum Mol Genet,2005,14(1):125-133.
[13]Risch N,de Leon D,Ozelius L,et al.Genetic analysis of idiopathic torsion dystonia in Ashkenazi Jews and their recent descent from a small founder population[J].Nat Genet,1995,9(2):152-159.
[14]Brassat D,Camuzat A,Vidailhet M,et al.Frequency of the DYT1 mutation in primary torsion dystonia without family history[J].Arch Neurol,2000,57(3):333-335.
[15]Grundmann K,Laubis-Herrmann U,Bauer I,et al.Frequency and phenotypic variability of the GAG deletion of the DYT1 gene in an unselected group of patients with dystonia[J].Arch Neurol,2003,60(9):1266-1270.
[16]Valente EM,Warner TT,Jarman PR,et al.The role of DYT1 in primary torsion dystonia in Europe[J].Brain,1998,121(Pt 12):2335-2339.
[17]Hjermind LE,Werdelin LM,Sorensen SA.Inherited and de novo mutations in sporadic cases of DYT1-dystonia[J].Eur J Hum Genet,2002,10(3):213-216.
[18]Major T,Svetel M,Romac S,et al.DYT1 mutation in primary torsion dystonia in a Serbian population[J].J Neurol,2001,248(11):940-943.
[19]Im JH,Ahn TB,Kim KB,et al.DYT1 mutation in Korean primary dystonia patients[J].Parkinsonism Relat Disord,2004,10(7):421-423.
[20]Matsumoto S,Nishimura M,Kaji R,et al.DYT1 mutation in Japanese patients with primary torsion dystonia[J].Neuroreport,2001,12(4):793-795.
[21]Cheng FB,Wan XH,Zhang Y,et al.TOR1A sequence variants and the association with early-onset primary dystonia in the Chinese Han population[J].Parkinsonism Relat Disord,2012,19(3):399-401.
[22]Lin YW,Chang HC,Chou YH,et al.DYT1 mutation in a cohort of Taiwanese primary dystonias[J].Parkinsonism Relat Disord,2006,12(1):15-19.
猜你喜欢
电子科技大学学报(2022年5期)2022-10-29
全科护理(2022年3期)2022-02-18
中国康复(2021年6期)2021-11-30
中国生殖健康(2020年4期)2021-01-18
中国生殖健康(2018年4期)2018-11-06
中国现代神经疾病杂志(2017年1期)2017-03-29
米娜·女性大世界(2016年8期)2016-08-17
Coco薇(2016年5期)2016-06-03
爆笑show(2015年9期)2015-10-24
湖北农业科学(2014年11期)2014-09-10
