
技术联盟组织间知识转移的类生物机制研究
2013-09-21 08:46张红兵,和金生,张素平
大连理工大学学报(社会科学版) 2013年4期
张 红 兵, 和 金 生, 张 素 平
(1.山西财经大学 管理科学与工程学院,山西 太原030031;2.天津大学 管理与经济学部,天津300072;3.加州大学 圣地亚哥分校医学院,美国 圣地亚哥92093)
知识是组织的战略核心资源[1],也是组织价值创造的主要来源[2]。组织要建立和保持自身的竞争力,须与外界进行交流,及时地获取和学习知识[3]。技术联盟为组织获取和学习外部知识提供了良好的机会,联盟内组织间高效的知识转移能够促进各方充分融合内外部知识,快速确立竞争优势[4]。随着联盟规模的不断扩大和内部成员组织结构的日益复杂,技术联盟组织间知识转移过程中所呈现的类生物特征引起了学术界的普遍关注,很多学者就这一现象进行了卓有成效的相关研究。Nardli、O’Day研究认为,知识转移与共享具有条件性、逐利性等生物特性[5];Holsapple、Singh则指出,组织与其他利益相关主体的知识交流情境形成了组织成长的生命周期,并以此构筑出组织的类生物系统[6];和金生、李江对组织知识发展中知识扩散与创造的类生物行为进行了解读[7];蒋天颖从自然生态角度勾勒出企业组织知识转移类生物形式的活动过程[8]。然而,这些研究均是从知识转移活动的外在条件、行为和形式出发进行研究,而对知识本身的生物属性却进行了“黑箱化”处理;其次,学者们的研究视角多聚焦于组织内部,对于组织间特别是技术联盟内组织间知识转移的活动规律却很少涉及。
基于此,本文借鉴仿生学原理对技术联盟组织间知识转移的类生物机制进行研究,力图在透析知识生物属性的基础上,探寻出技术联盟组织间知识转移活动的发生发展来源及具体实现过程。
一、知识的生物属性
知识属性的研究一直是科学哲学和科学社会学等学科的重点研究内容。无论是科学社会学代表人物默顿提出的“求真”的标准解释,还是后来爱丁堡学派的“强纲领”[9],都是仅从知识活动的外在条件出发来归纳知识的特征和演进规律,无一例外的未能打开知识属性这一“黑匣子”。当代著名哲学家波普尔认为,知识活动同生物进化有着惊人的相似。中国最早研究知识基因的学者刘植惠指出,知识与生物同属于发达的演化系统,所有发达的演化系统的发育必定与遗传基因控制机理相同[10]。本文将知识活动过程中所表现的各种特征与生物的生存、繁衍过程进行类比,发现知识具有下述生物属性:
(1)延续性。生物广泛存在于整个世界,种类繁多,形态和性质各异,但生物种群有自身的起源,其发展、进化和演绎过程都表现出稳定的延续性。知识也是种类繁多,内容各有不同,其发展也具有相应的延续性:无论任何知识都必然会有一些起因线索,现有知识只能在过去的知识基础上演化而来。知识的延续性特征使得人们只能渐进学习,只能吸收、消化可与原有知识基础相连接的新知识,不能拔苗助长也不能跨越式学习。虽然人们也会接触到自己难以接受的知识,就像给没有任何数学基础的人讲一堂微积分,他所接受的只能是该听者原有知识相关联的部分内容。
(2)遗传性。生物亲代性状通过遗传传递给子代,亲代生物的性状在子代中得以体现,子代再次将其性状传递给其后代,如此代代相承使得生物性状保持着一定的不变性。如同生物性状一样,人类的一些特有知识如经验、真理等在相当长的一段时间内亦能够经受住考验,其独特性状和核心结构稳定不变,具有明显的遗传性。
(3)变异性。生物必须能够适应不断变化的环境才能得以生存,否则会被淘汰。经过一代又一代的进化,生物适应环境的性状被保留下来,而其他性状特征则逐渐消亡,这种性状特征的渐变就被称为生物的变异性[11]。非常相似的是,人类对自身与外界的认识也是不断变化的,认识过程往往蕴含着痛苦的思辨、反复的争论,就像生物接受环境选择一样只有符合客观事实或规律的认识才能得以传承[12],显然知识(认识)经过变异后进一步获得了顽强的生命力。
(4)群合性。生物个体为了增加生存几率,往往相互支持和依赖,自组织形成一个个群落,呈现出不同程度的群合性。同样,知识发展的主要形式如跨越组织边界的知识交流、各类学科知识的碰撞等使得知识活动体现出了大量的群体合作特征——群合性。
(5)酶合性。生化反应中为了增加作用效果或速率,往往会加入一种生物酶进行催化。没有生物酶的酶合作用,生化反应一般很难完成。同样,知识的活动越来越复杂,人类社会的专业分工愈来愈细,导致现代知识活动根本不可能离开各种媒介的支持(知识发展需要酶合)。知识发展中的酶合作用,正如学生学习知识过程中导师的“传道授业解惑”。人类社会的发展、组织的生存都越来越离不开各类媒介的推动。
(6)环境依存性。任何生物离开了现有的环境如土壤、水、空气等,将不能够存活。相应的,知识发展也需要适宜的社会、经济、政治以及文化等环境。
二、技术联盟组织间知识转移的类生物机制模型
有了以上知识生物属性的论述,一个有关知识活动的过程跃然纸上:知识,是一种具有“环境依存性”的有机生长生物,其“遗传”和“变异”发展总是起自一种“延续”的初始知识,在受到一定刺激物催化“酶合”时,通过一定的途径吸收外界知识与自身已有知识在“群合”场所内进行融合、重构,从而产生处理问题的新方案、新主张等新知识,这种初始知识的传承或变异后的新知识产生都是知识转移成功的表现。由此,本文将重新思考技术联盟组织间知识转移活动的内在规律,类比生物发酵的组成要素[13],构建出一个包括知识菌株、知识母体、知识酶、知识环境、知识技术、发酵吧和新知识等7种发酵要素的技术联盟组织间知识转移的类生物机制模型,如图1所示。
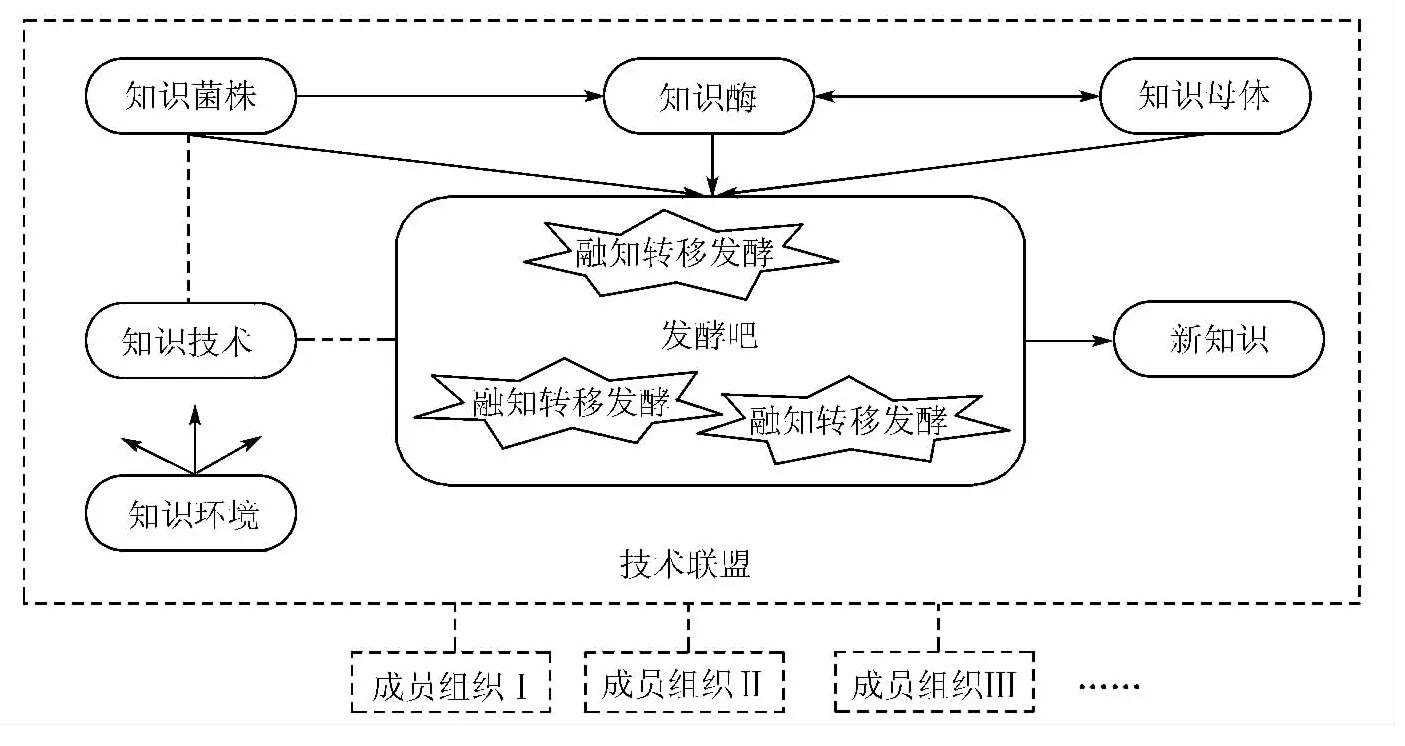
图1 技术联盟组织间知识转移的类生物机制模型
(1)知识菌株。任何知识活动都具有一定的目的性,如果没有一定的目的,人类便不会开始进行知识活动。因此,知识转移起源总是来自一种“初始的知识缺口”,本模型称之为知识菌株。在技术联盟中,知识菌株主要表现为各成员组织间的知识“势差”。技术联盟内的知识分布具有非均衡性,各成员组织在知识深度和广度方面拥有的知识存量各不相同,在知识区位中有着高低位势的差别,存在着相应的知识“势差”。技术联盟中知识“势差”的存在,使得知识不断从高位势成员组织向低位势成员组织进行转移,低位势成员组织将所吸收知识与自身已有知识融合、变异后产生新知识,进而使得联盟中的知识“势差”在时间维度上不断演变,各方所拥有的知识“势差”不断降低,当“势差”降低到零时,成员组织停止吸收知识,技术联盟整体不再产生知识转移活动。由此,知识“势差”作为关键知识菌株,是技术联盟组织间知识转移的前提。需要指出的是,联盟内成员组织间高位势和低位势属性是相对的,成员组织与其他伙伴相比在某一类知识上处于高位势却可能在其他类知识上处于低位势。
(2)知识母体。“知识母体”是一个过程性概念,专指学习者已经掌握并参与学习过程的知识集合,包括技术联盟内各成员组织的基础知识和专业知识。基础知识是相互间交流的通用语言,联盟双方较高关联性的基础知识是知识转移的必要条件,如果联盟双方的基础知识的关联性较小,就会导致接收方不能将准确的理解联盟内知识源发送过来的知识,无法将自身已有知识与发送过来的知识进行有效类比、匹配和融合[14]。拥有优势知识资源的企业组织向联盟伙伴转移复杂的知识时,须提前将一些简单配套的知识转移给接收方,否则将影响后续的知识转移效果。与基础知识相比,专业知识是对特定技术领域深入的、个性化的理解,其差异性是联盟内成员组织间知识转移的触发器。若成员组织间专业知识的差异性缩减,将导致相互之间可供转移的知识资源变少,极端情况下如果双方拥有完全一致的知识资源,组织间就不可能产生知识转移活动。
(3)知识酶。从生化反应的原理得知,酶分子通过某种耦合作用,可以有效地激发反应物的活化能,加速生化反应的完成。在技术联盟组织间的知识转移过程中,知识酶是指有助于知识转移顺利进行的各种因素,它的主要任务是削弱或消除成员组织之间存在的各种各样阻碍知识扩散的障碍,主要包括成员组织间信任与承诺。成员组织间信任是联盟成员对伙伴不采取机会主义行为且有能力实现其义务的预期,在信任的基础上,联盟成员更愿意分享知识资源,会更大程度地提高知识转移的强度,尤其是隐性知识转移只能通过技术联盟内信任的社会网络进行[15];承诺是联盟内相关决策者在知识转移合作中资源的投入意愿,成员组织间的承诺是维持、强化及加深伙伴间知识转移的有效途径。此外,内生与外附的激励机制等亦是一种有效的知识酶,在知识转移过程中具有显著的催化作用[16]。
(4)知识环境。知识转移脱离不了外部环境,类似各种外部环境如气压、湿度、温度等对微生物能否成功发酵所起的重要作用一样,技术联盟组织间成功的知识转移也需要适宜的知识环境。参与联盟合作的企业组织均面临着复杂的外部环境,这种环境的复杂性限制了企业组织获取外部优势知识资源的能力,极大地增加了组织间知识转移失败的风险。因此,盟主企业应积极地采取各种措施营造有利于成员组织间知识转移的外部环境,如鼓励创新的科技政策、浓郁的知识共享文化、容忍创新出错的氛围等。
(5)知识技术。这里指促进知识转移的科学与信息技术。Ethernet、Internet、Intranet、Extranet网络,Web2.0信息技术,Groupware群件,Database warehouse,知识仓库等科学技术的迅猛发展,使得成员组织间的实体与虚拟空间实时交流成为可能。尤其是知识仓库的发展,不仅能像以往一样对分散于成员组织内部零散的显性知识进行编码化处理,还可以对不便于格式化、不易交流的隐藏在联盟内成员组织个体人员大脑中的隐性知识从重要性、获取难易程度、可共享性、独有性等方面进行科学分类和系统测度,实现成员组织隐性知识的有序化、编码化,从而为提高知识转移效率提供有力支撑。
(6)发酵吧。“发酵吧”既是物质空间(各种物理办公场所)与虚拟的超物质空间(通过无线网络实现的远程PtoP视频聊天室、3D空间沟通),也是精神空间(共享的经验、观念和理想等),或者是这三者的组合。发酵吧为知识菌株、知识母体、知识酶等各种要素“群合”提供了场所,知识管理者必须在技术联盟内依据现实条件设立对应类型的发酵吧。
(7)新知识。各发酵要素在“发酵吧”内发酵后,本文称之为“融知转移发酵”,知识源有价值的知识被转移至接收方组织,所转移的知识可能原封不动成为接收方组织累加的新知识,也可能与接收方组织原有的互补性知识融合后变异为全新的新知识。这种累加或全新的新知识的出现,均表明技术联盟组织间知识转移得以成功实施。
图4为明显异常图例,可见地下12ns白色方框处,有非常明显的空隙,同相轴扭曲或断裂,判断该处异常为疑似脱空区。
在图1所示的模型中,各发酵要素经过“融知转移发酵”来有效地实现技术联盟组织间的知识转移。“融知转移发酵”作为技术联盟组织间知识转移的重要组成部分,其运行过程紧紧地围绕着知识重构和知识扩散两个环节进行。
知识重构是指技术联盟内成员组织间相互沟通、学习后,不同知识的交叉、碰撞导致原有知识基因的改变,从而催生出一个新的方案、创意或理论等新知识的创造过程。知识的遗传性与变异性在此过程中同时体现:一方面联盟内单个成员组织的知识经过复制原封不动成为各个接收方组织的累加知识,知识得以遗传;另一方面由于参与交流的各方知识背景的不同对“共享知识”产生不同的理解,进而触发各方已有知识与“共享知识”匹配、融合、成长后转变为新的知识,知识得以变异。知识重构是为了解决各联盟成员面临的尖锐的问题和矛盾,更加注重辩证的思考方式,力求在更高层次上实现新的统一,其参与主体的范围一般较小,主要来源于成员组织的相关决策者或管理者。开放、自由的知识环境、鼓励不同思想之间进行碰撞的知识酶、快速发展的知识技术等发酵要素均能激发知识重构的运行。知识重构所产生的新知识,会使得诸如知识菌株(成员组织间的知识势差)、知识酶(如知识转移意愿)等发酵要素的特征发生变化,由此将引发技术联盟内更大范围的组织间协同学习,即知识扩散。
知识扩散是将显性或隐性知识进行传播、渗透和应用的组织间协同学习过程。知识扩散在技术联盟内广泛存在,有的知识是在大范围内扩散,有的知识仅仅在相临的两个成员组织间扩散。技术联盟中若一个成员组织的知识发生了改变,会导致与之相邻的相关知识领域的合作伙伴之间知识势差、知识需求等发生改变,由此激发、推动该领域相关知识产生剧烈的扩散过程。即使参与联盟的各成员组织可能不在同一个地区,若上游成员组织技术发生更新(例如,有了新材料的产生和批量使用),下游成员组织就会进行相应的技术变革和知识更新;同样,下游成员组织若提出了新的产品设计,其配件的设计变化就会依次向上游进行传递。联盟内各成员组织的研发很少独自进行,往往要上下游共同参与到新产品或新技术的研究、交流中来,通过协同形式的学习降低新知识的扩散周期。
知识重构和知识扩散过程中,起联接作用的发酵要素的培育至关重要。例如,对知识菌株进行培育——加强技术联盟中专业研究机构与企业的合作程度,可以实现创意与实施的完美结合,像清华大学与日本三菱公司进行的膜生物反应器的项目合作,虽然都有突出的技术专长,但前者更多优势是无限的创意设计,后者是敏锐的市场应用实践。
知识重构和知识扩散构成了发酵吧内的“融知转移发酵”过程(见图2),其中任何一个环节出现问题,都会导致“融知转移发酵”受阻,使得技术联盟组织间 知识转移能力降低甚至失效。
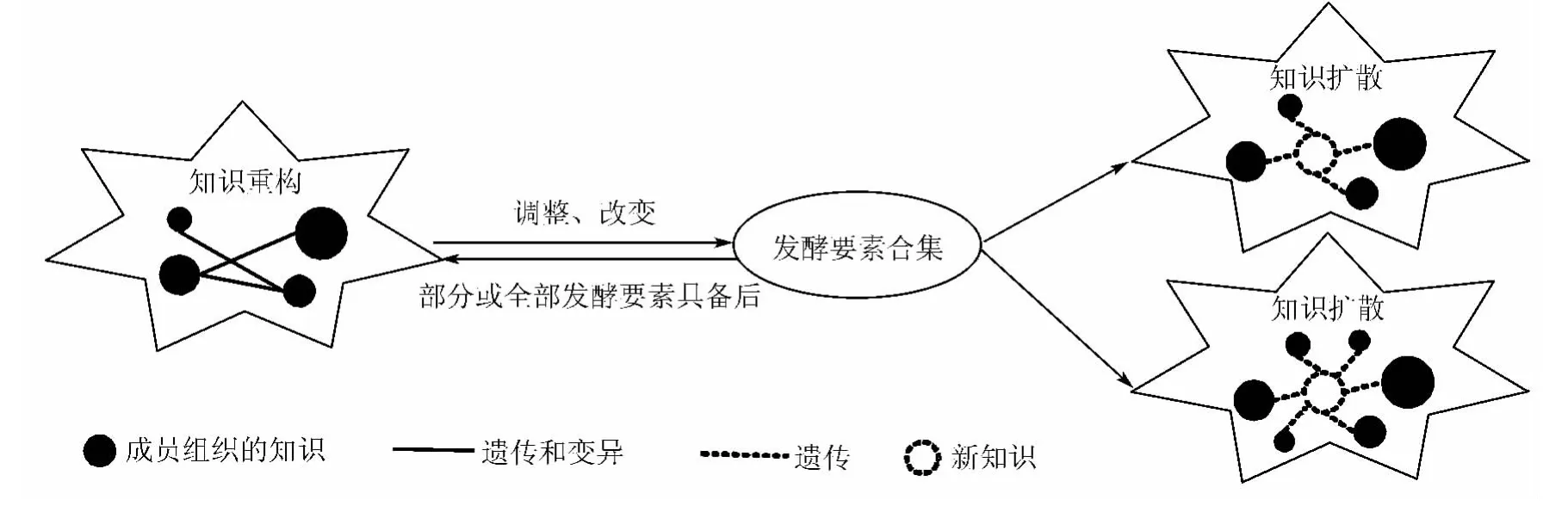
图2 “融知转移发酵”过程
三、案例分析
技术联盟作为当前一种快速发展的组织合作模式,其组织间知识转移案例比比皆是,本文就对中国3G技术联盟组织间知识转移的案例进行分析。
1999年3月,在科技部、信息产业部、国家计委等部委组织协调下,由中国移动、中国联通、北京大学、清华大学、大唐电信和深圳华为等国内20多家高校、研究所和企业近3000名科技人员合作组成“中国第三代移动通信系统技术联盟”,即中国3G技术联盟。该联盟自成立以来一直以高效的知识转移与创新能力著称,已成功将分散在“产、学、研”各组织的局部知识资源优势整合成了我国移动通信产业的整体优势,开发了包括无线接入网、核心网、终端现场实验系统等近百项具有自主知识产权的新产品。中国3G技术联盟高效的知识转移能力来源除了其合理的联盟伙伴组合、联盟内有效的利益协调与激励机制、鼓励相互学习的文化与政策环境、发达的信息通讯系统外,还有一个非常富有成效的管理制度——例行的专题会议。
例行的专题会议是针对具体研究专题每周举行的会议。比如,中国3G技术联盟在进行“第三套试验系统”(指在车载环境下提供速率达144kb/s的数据传能力,并能提供上网浏览和移动视频多媒体业务的技术体系)研发过程中,每个星期一将清华、北大、大唐电信等相关单位的科研主管召集到一起,讨论各单位在上周“第三套试验系统”技术产品原形协作开发中出现的问题以及科研负责人的意见、希望、应对策略,从而调整和决定各单位下一步开发的方针与方向。星期二举行参与人员扩展到各单位技术骨干、每个子专题科研成员的研讨会议,上午传达前一天会议中所讨论的方针和协作开发策略,下午按照不同的子专题分别举行小组会议,针对上午所涉及的相关问题点进行详细讨论。两天例行的专题会议结束之后,参会人员将所得信息充分传递到各自所在单位,在最短的时间内应用于工作实践。
从上述案例可以看出,中国3G技术联盟组织间知识转移所需的发酵要素——知识菌株(知识资源优势不同的高校、研究所和企业之间存在的知识势差)、知识母体(各单位科研主管、技术骨干以及科研成员的知识集合)、知识酶(联盟内有效的利益协调与激励机制)、知识环境(鼓励相互学习的文化与政策环境)、知识技术(发达的信息通讯系统)、发酵吧(例行的专题会议)均已形成,且各发酵要素在发酵吧内“融知转移发酵”过程的重构和扩散两个环节也非常清晰:星期一召集各单位科研主管的专题会议,经过讨论和思考,形成下一步技术和产品开发的方针、方向、策略,这是一个知识的重构过程,将导致发酵要素合集的特征产生调整、改变(如新知识的出现,知识酶的催化效应增减等);而第二天上午的会议,通过传达这些方针、方向和策略形成知识在成员组织间扩散,在下午的分组会议中,参会者根据各自所面对的情况进一步对知识进行消化,形成具体可实施的方案,最后扩散至每一个成员组织内部。
四、结 论
通过前文关于技术联盟组织间知识转移的类生物机制的理论剖析,并结合中国3G技术联盟组织间知识转移案例的探讨,本文得出如下结论:
第一,知识活动与生物的繁衍、生存过程非常相似,具有6种典型的生物属性,即延续性、遗传性、变异性、群合性、媒合性和环境依存性。以往也有学者采用仿生学方法对知识活动进行分析,然而研究成果均是从知识转移活动的外在条件、行为和形式出发进行辨别和阐释,对此Reydon等在综述仿生学发展历程中指出,很多“仿生学对具体管理实践无较大相关性”的批评积聚在研究单位的选择上,并建议研究单位应该偏向于客体对象(技术联盟知识转移领域的客体对象是指“知识”)[17]。本文遵循Reydon的仿生学观点,类比生物的发展过程明确了本研究客体对象——知识的主要生物属性,在一定程度上对回应关于仿生学的批评做出一些贡献。
第二,技术联盟组织间知识转移活动的发生发展来源于联盟内出现的知识菌株,同时也离不开知识母体、知识酶、知识环境、知识技术、发酵吧等发酵要素的支撑和协作。知识菌株是知识转移活动产生的前提条件,联盟内的成员组织只有在识别自身知识缺口,产生知识菌株(感受到与知识源之间的知识势差)之后才会触发知识转移需求;进一步,在其他各发酵要素的推动下,知识转移活动方能逐步平稳进行。
第三,各发酵要素经过“融知转移发酵”来实现技术联盟组织间的知识转移,其中知识重构和知识扩散构成了“融知转移发酵”的全部过程,无论是知识重构或知识扩散环节出现问题均会对知识转移造成影响。对此,要确保知识重构和知识扩散的“融知转移发酵”过程顺利进行,起联接作用的各发酵要素如知识菌株、知识母体、知识酶等的培育至关重要,这也是提升技术联盟组织间知识转移能力的具体实践途径。
然而,应当指出的是,本文的研究依然存在着一定程度的局限性,需要在今后逐步的完善和丰富。如,不同性质技术联盟(竞争性联盟与非竞争性联盟、股权式联盟与契约式联盟、渐近性创新联盟与突破性创新联盟等)的发酵要素内部构成异同,各发酵要素在知识转移不同时段相互间关系的非均衡性、动态性等,都是未来值得进行深入研究的议题。
[1]NONAKA I.A dynamic theory of organizational knowledge creation[J].Organization Science,1994,5(1):14-37.
[2]LEWIS D J,EDWIN H N,BOHUMIR P.Knowledge,in novation and share value[J].International Journal of Management Reviews,2002,(2):101-134.
[3]NICOLAI J F,KENNETH H,SNEJINA M.Governing knowledge sharing in organizations:levels of analysis,governance mechanisms,and research directions[J].Journal of Management Studies,2010,47(3):455-482.
[4]JIANG Z S,HU L Y,CHEN K K.Decisions of knowledge transfer in technology innovation alliance:a stackelberg leader-followers model[J].Operational Research,2010,(2):231-242.
[5]NARDLI B A,O’DAY V L.Information Ecologies:Using Technology With Heart[M].Massachusetts:The MIT Press,1999.102-188.
[6]HOLSAPPLE C W,SINGH M.The knowledge chain model:activities for competitiveness[J].Expert System with Applications,2001,20(1):77-98.
[7]和金生,李江.知识发展的类生物模型[J].科学学研究,2008,26(4):679-684.
[8]蒋天颖,程聪.企业知识转移生态学模型[J].科研管理,2012,33(2):130-137.
[9]浦根祥.知识属性、动因及增长方式的演变 [J].科学学研究,2000,18(1):29-34.
[10]刘植惠.知识基因理论研究[J].重庆图情研究,2003,(13):4-10.
[11]STEIBEL,JUAN P,SUCHYTA,et al.Tackling high variability in gene expression studies[J].Genomics &Proteomics,2005,(5):30-32.
[12]唐建生,和金生.基于知识发酵理论的组织知识共享问题研究[J].科技管理研究,2007,(5):230-233.
[13]LIN Z,LI W H.Expansion of hexose transporter genes was associated with the evolution of aerobic fermentation in yeasts[J].Molecular Biology and Evolution,2011,28(1):131-142.
[14]徐小三,赵顺龙.知识基础互补性对技术联盟的形成和伙伴选择的影响[J].科学学与科学技术管理,2010,(3):101-106.
[15]OXLEY J,WADA T.Alliance structure and the scope of knowledge transfer:evidence from U.S.-Japan agreements[J].Management Science,2009,55(4):635-649.
[16]王永强,朱方伟,蒋兵.技术引进中研发人员隐性技术知识转化激励研究[J].大连理工大学学报(社会科学版),2011,32(1):31-35.
[17]REYDON T A C,SCHOLZ M.Why organizational ecology is not a darwinian research program[J].Philosopy of the Social Sciences,2009,39(3):408-439.
猜你喜欢
城市建设理论研究(电子版)(2022年27期)2022-09-30
城市建设理论研究(电子版)(2022年10期)2022-06-08
城市建设理论研究(电子版)(2022年4期)2022-06-08
城市建设理论研究(电子版)(2022年9期)2022-06-07
哈尔滨工业大学学报(2022年5期)2022-04-19
草地学报(2022年3期)2022-03-28
摄影世界(2022年1期)2022-01-21
今日农业(2021年11期)2021-11-27
知识经济·中国直销(2018年12期)2018-12-29
商周刊(2017年6期)2017-08-22
