
酿酒酵母GSH I基因的克隆、表达与结构分析
2013-12-06 07:14宋冬梅朱益波周丽丽郑丽雪
食品工业科技 2013年13期
宋冬梅,朱益波,周丽丽,郑丽雪,齐 斌,*
(1.吉林农业大学,食品科学与工程学院,吉林长春130118;2.常熟理工学院发酵工程技术研究中心,苏州食品生物技术重点实验室,江苏常熟215500)
谷胱甘肽(glutathione,GSH)是在γ-谷氨酰半胱氨酸合成酶(GSH I)和谷胱甘肽合成酶(GSHⅡ)的连续催化下合成的含巯基活性三肽,具有解毒、抗氧化和抗衰老等重要的生理功能[1-3],在食品、医药和生物等领域有着广泛地应用[4-7]。但是我国GSH的生产满足不了市场的需求。近年来,GSH生产研究的热点是利用DNA重组技术构建高产GSH的重组菌。Murata等[8]构建了具有较高GSH催化活性的重组 Escherichia coli。沈立新等[9]构建了含 GSH I及GSHⅡ基因的质粒pTrc-gsh,并在E.coli中得到了活性表达。Liao等[10]构建的E.coli GSH I和GSH Ⅱ基因融合表达质粒,提高了胞内GSH含量。这些研究在一定程度上提高了重组菌GSH的产量,但总的来说都不理想。GSH I是GSH合成途径的关键酶和限速酶[11],构建高GSH I活性的菌株将有助于提高重组菌GSH的产量。本文从实验室保存的一株高产GSH的Saccharomyces cerevisiae 2-10515出发,采用PCR技术扩增得到GSH I基因,并在E.coli中实现表达,同时利用生物信息学方法对GSH I基因序列和结构进行分析和预测,为进一步提高GSH I表达量和活性提供理论依据。
1 材料与方法
1.1 材料与仪器
S.cerevisiae 2-10515、E.coli JM109、E.coli BL21(DE3)、质粒pET-28a 本实验室保藏;DNA限制性内切酶Sac I和HindⅢ、Taq DNA聚合酶、T4 DNA Ligase、pMD19-T 载体、DNA Marker、质粒小量提取试剂盒、DNA纯化及胶回收试剂盒等 TaKaRa公司;其他试剂 均为国产分析纯;YEPD培养基、LB培养基 配方见文献[12]。
高速台式离心机Legend Micro 17R Thermo公司;PCR仪、Gel Doc XR、水平电泳系统、垂直电泳系统、凝胶电泳成像仪 Bio-RAD公司;恒温金属浴Eppendorf公司;SW-CJ-ZF净化工作台 苏州净化设备有限公司;恒温培养箱 太仓市华美生化仪器厂;电子天平 Mettler公司。
1.2 实验方法
1.2.1 引物设计 根据Gene bank里已公布的S.cerevisiae S288c的gsh I基因设计引物,分别在上游和下游引物中引入酶切位点Sac I和HindⅢ:
P1(Forward):CACGAGCTCATGGGACTCTTAGCTTT酶切位点Sac I
P2(Reverse):CCCAAGCTTTTAACATTTGCTTTCTA酶切位点HindⅢ
1.2.2 克隆载体的构建 采用周小玲等人[13]的方法提取S.cerevisiae 2-10515的基因组 DNA,以其为模板,P1和P2为引物进行PCR扩增,得到gsh I基因。PCR 反应参数为:95℃ 5min;94℃ 30s,58℃ 30s,72℃ 2min(30cycles);72℃ 10min。将gsh I连接到克隆载体pMD19-T上,构建克隆质粒 pMD19-T-gsh I,并转化E.coli JM109,得到E.coli JM109(pMD19-T-gsh I),提取克隆质粒进行酶切验证,送上海生工生物有限公司测序。
1.2.3 E.coli重组表达载体的构建 分别用限制性内切酶Sac I和HindⅢ酶切克隆质粒pMD19-T-gsh I和质粒pET-28a,酶切产物经胶回收纯化,16℃连接过夜,转化E.coli DH5α。抽提重组质粒,经酶切验证后测序。
将重组表达质粒 pET-28a-gsh I热激转化到E.coli BL21(DE3)。
1.2.4 GSH I的诱导表达 挑取单菌落接入LB培养基(含100mg/mL Amp),37℃培养至 OD550为 0.5左右时,在34℃,IPTG(0.1mmol/L)条件下,诱导表达。
1.2.5 SDS-PAGE检测 按《分子克隆》[14]操作说明进行SDS-PAGE电泳后,考马斯亮蓝R-250染色,脱色过夜,观察结果。
1.2.6 GSH I酶活性的测定 取5mL培养液12000×g离心5min,弃上清,用50mmol/L,pH7.0的磷酸缓冲液洗涤,离心后收集菌体。用同样的磷酸缓冲液4mL悬浮菌体后超声波破碎[15],加入GSH前体反应液(谷氨酸、甘氨酸和半胱氨酸浓度均为40mmol/L,MgCl2和ATP浓度均为50mmol/L,GSH I浓度为2g/L)4mL,37℃反应 30min,沸水浴 2min以终止反应,用ALLOXAN试剂法[16]测GSH的生成量。酶活力单位(U)定义为37℃,pH7.0条件下,每分钟催化合成1μg GSH所需的酶量。
1.2.7 GSH I的结构分析 利用DNAMAN软件推导出目的基因的氨基酸序列,分析其与Gene bank数据库中其它物种同源蛋白氨基酸序列的一致性。
通过蛋白分析专家系统(ExPASy,http://ca.expasy.org/)提供的分析工具分别预测蛋白质的理化性质、一级结构和二级结构。
利用3D-JIGSAW模拟GSH I的三级结构。
2 结果与分析
2.1 基因克隆
以S.cerevisiae 2-10515基因组DNA为模板进行PCR扩增反应,扩增产物纯化后电泳结果见图1。图1显示:目的条带在2000bp附近,与预计的条带大小相吻合。

图1 gsh I基因的PCR纯化产物Fig.1 PCR purification product of gsh I
2.2 重组表达载体的构建
成功构建的表达载体pET-28a-gsh I的结构示意图如图2。将其用Sac I和HindⅢ酶切鉴定,结果如图3。经酶切的重组质粒含有单一的约2Kb插入片段,这表明gsh I基因已插入质粒pET-28a中。测序结果证明阅读框插入位点正确。

图2 重组表达载体pET-28a-gsh I结构示意图Fig.2 Construction of recombinant expression plasmid pET-28a-gsh I simple

图3 重组质粒pET-28a-gsh I/Sac I+HindⅢ酶切鉴定Fig.3 pET-28a-gsh I digested by Sac I and Hind Ⅲ
2.3 SDS-PAGE电泳分析
SDS-PAGE电泳后考马斯亮蓝染色,脱色过夜,观察结果如图4所示:2、3、4泳道与阴性对照(1泳道)相比,在81Ku附近有一条明显的蛋白条带,除去载体上的his-tag(分子量约为3ku),其实际的蛋白质分子量约为78ku,与预测的GSH I蛋白大小相符,很可能为表达的GSH I蛋白,该蛋白以融合蛋白的形式存在。

图4 SDS-PAGE分析图谱Fig.4 SDS-PAGE analysis of expression products
2.4 酶活力的测定
重组菌的GSH I活力测定结果见图5,34℃诱导4h时,其活力达到46.09U/mg湿菌,随着诱导时间的延长,GSH I活力并没有增加,最佳诱导时间为4h。优化各种表达条件,酶活力无明显改善。

图5 重组菌GSH I酶活力曲线Fig.5 GSH I enzymatic activity curve of recombinant strain
2.5 GSH I基因的结构分析
2.5.1 Blast分析结果 该基因全长为2037bp,编码678个氨基酸。对GSH I氨基酸序列进行同源多重序列比对(如图6)发现:序列中部为保守序列,其关键氨基酸都在该保守序列中;与Gene bank中S.cerevisiae S288c、Macaca mulatta、Musmusculus、Branchiostoma floridae同源基因的氨基酸序列一致性分别为99%、42%、42%、41%。

图6 GSH I的Clustal多重序列比对Fig.6 Multiple alignment of GSH I by Clustal
2.5.2 GSH I蛋白质的理化性质 利用ExPASy中ProtParam预测GSH I的理论分子量为78281.6u;等电点为5.87;含有9个半胱氨酸;其中第164位和第642位半胱氨酸有形成二硫键的可能性;该蛋白质的性质不稳定,在溶液中的不稳定指数为42.19,高于阈值40;脂溶性指数为77.52;亲水指数(GRAVY)为-0.507,为亲水性蛋白。其成熟肽N端为蛋氨酸时,在哺乳动物网状红细胞体外表达的半衰期为30h,在酵母和大肠杆菌中表达的半衰期分别为20、10h。
2.5.3 GSH I的二级结构 利用ExPASy中PredictProtein分析蛋白的拓扑结构和二级结构,该蛋白没有信号肽,为非分泌蛋白;在253~272aa、285~308aa和484~511aa处有三个跨膜区;α-螺旋∶延伸链:β-折叠∶无规则卷曲 =35.10∶16.52∶8.85∶39.53,表明蛋白质二级结构的主要结构元件是α-螺旋和无规则卷曲,具有较多的无规则卷曲,不利于其稳定;mRNA的二级结构在起始密码子处形成发卡结构。
2.5.4 GSH I的三级结构 将GSH I的氨基酸序列提交到3D-JIGSAW服务器预测GSH I三级结构,用Pymol软件打开,如图7所示,发现酶结构较松散,侧链间相互作用不强,这与ProtParam的预测结果相一致。
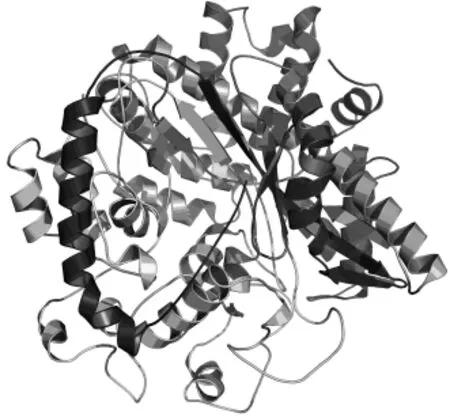
图7 GSH I三级结构图Fig.7 3D structure of GSH I
3 结论与讨论
已有学者在S.cerevisiae和Pichia pastoris中表达了 GSH I[17-18],但这些报道中 GSH I 的表达量不高,酶活力也较低,使得构建好的菌株无法达到工业化生产的要求。S.cerevisiae是高产GSH的菌株,将其GSH I在E.coli中表达,有望构建高活性的GSH I重组菌,为工业化生产GSH奠定基础。本研究成功克隆了S.cerevisiae 2-10515的gsh I基因,将编码GSH I的基因定向克隆到pET-28a载体上,构建了重组菌株 E.coli BL21(DE3)/pET-28a-gsh I,34℃ 诱导 4h时,活力达到 46.09U/mg 湿菌,高于 Fan[17]、饶志明[18]等构建的重组菌株。
利用生物信息学方法分析S.cerevisiae 2-10515 GSH I序列可知:a.gsh I基因转录出的mRNA在5'端起始部位区形成了发卡结构,有研究证实这种发卡结构会遮蔽SD序列和起始密码子部位能阻止30s核糖体亚基的结合,从而减慢翻译起始频率[19],降低相应蛋白质的表达量。B.GSH I在溶液中的不稳定指数为 42.19,高于阈值 40;GSH I亲水指数为-0.507,为亲水性蛋白;该酶分子中含有较多的无规则卷曲,结构比较松散。这些都说明GSH I稳定性较差[20],不利于活性的提高。所以,从基因水平对S.cerevisiae 2-10515进行分子改造能进一步提高该酶的表达量和活性。
Huang[21]通过对 mRNA 结构的改造,消除了起始密码子附近mRNA的二级结构,从而提高了木聚糖酶的表达量;Jeong[22]、Mahadevan[23]等通过同源建模及结构分析,利用定点突变增加了酶的稳定性。这些研究都为后续对S.cerevisiae GSH I分子改造提供了很好的基础。本文在 E.coli中成功表达了S.cerevisiae来源的GSH I,为工业应用奠定了基础;并对其进行了初步的生物信息学分析,为后续研究指明了方向。
[1]Grill E,Löffler S,Winnacker E L,et al.Phytochelatins,the heavy metal binding peptides of plants,are synthesized from glutathionebyaspecific γ-glutamylcysteine dipeptidyl transpeptid-adse(phytochelatin synthase)[J].Proc Natl Acad Sci USA,1989,86:6838-6840.
[2] MeisterA.Glutathione metabolism and its selective modification[J].Biol Chem,1988,263:27205-27208.
[3]Sanchez-Femandez R,Fricker M,Corben L B,et al.Cell proliferation and hair tip growth in the Arabidopsis root are under mechanistically different forms of redox control[J].Proc Natl Acad Sci USA,1997,94:2745-2750.
[4]杨培慧,齐剑英.谷胱甘肽的应用及其检测方法[J].中国生化药物杂志,2002,23(1):52-54.
[5]冮洁,单立峰.谷胱甘肽的制备及其应用[J].饲料工业,2007,28(15):15-17.
[6]周宇光,付国平,肖仔军.谷胱甘肽的生产和应用研究进展[J].食品工业科技,2003,24(3):89-91.
[7]Spector D,Laban J,Toledanom B.A genetic investigation of the essential role of glutathione[J].Biol Chem,2001,276(10):7011-7016.
[8]Murata K,Miya T,Gushima H,et al.Cloning and amplification of a gene for glutathione synthetase in E.coli B[J].A gric Biol Chem,1982,47:1381-1383.
[9]沈立新,魏东芝,赵哲峰,等.谷胱甘肽合成酶系的克隆、测序与表达[J].生物工程学报:2001,17(1):98-100.
[10]Liao X Y,Shen W,Chen J.Improved glutathione production by gene expression in Escherichia coli[J].Letters in Applied Microbiology,2006,43:211-214.
[11]Ohtake Y,Watanabe K,Tezuka H,et al.Expression of glutathione synthetase gene of Escherichia coli B in Saccharomyces cerevisiae[J].J Ferment Bioeng,1989,68(6):390-394.
[12]黄培堂 .分子克隆实验指南[M].北京:科学出版社,2005.
[13]周小玲,沈微,饶志明,等.一种快速提取真菌染色体DNA 的方法[J].微生物学通报,2004,31(4):89-92.
[14]Sambrook J,E F Fritsch,T Maniatis.Molecular cloning:A laboratory manual(Second edition)[M].NY:Cold spring harbor laboratory,1989.
[15]沈立新,魏东芝,赵哲峰,等.谷胱甘肽合成酶在大肠杆菌中的高效表达及性质[J].华东理工大学学报,2000,26(4):342-345.
[16]李从军,谢爱娣.酵母提取液中谷胱甘肽定量测定方法的比较[J].食品发酵工业,2010,36(7):158-162.
[17]Fan XY,He XP,Guo XN,et al.Increasing glutathione formation by functional expressing of the γ-glutamylcysteine synthetase gene in Saccharomyces cerevisiae[J].Biotechnol Lett,2004,26(5):415-417.
[18]饶志明,艾丽静,沈微,等.gsh I基因的克隆及其在巴斯德毕赤氏酵母中的表达[J].应用与环境生物学报,2007,13(2):257-260.
[19]Cruz-Vera L R,Magos-Castro M A,Zamora-Romo E,et al.Ribosome stalling and peptidyltRNA drop- off during translational delay at AGA codons[J].Nucleic Acids Res,2004,32(15):4462-4468.
[20]王大成.蛋白质工程[M].北京:化学工业出版社,2002.
[21]Huang H Q,Yang P L,Luo H Y,et al.Hingh-level expressin of a truncated 1,3-1,4-β-D-glucanase from Fibrobacter succinogenes in Pichia pastoris by optimization of codons and fermentation[J].Appl Microbiol Biotechnol,2008,78(1):95-103.
[22]Jeong M Y,Kim S,Yun C W,et al.Engineering a denovo internal disulfide bridge to improve the thermal stability of xylanase from Bacillus stearothermophilus No.236[J] .J Biotechnol,2007,127(2):300-309.
[23]Mahadevan S A,Wi S G,Lee D S,et al.Site-directed mutagenesisand CBM engineering ofCel5A(Thermotoga maritima)[J].FEMS Microbiol Lett,2008(2):205-211.
猜你喜欢
环球时报(2022-09-20)2022-09-20
中国医学影像学杂志(2021年6期)2021-08-13
今日农业(2020年24期)2020-12-15
食品科学(2018年10期)2018-05-23
水产科学(2016年1期)2016-12-19
天然产物研究与开发(2016年6期)2016-06-05
兽医导刊(2016年12期)2016-05-17
中国继续医学教育(2015年3期)2016-01-06
西南医科大学学报(2015年1期)2015-08-22
中国当代医药(2015年9期)2015-03-01
