
条件性的关联学习和消退过程的性别差异*
2014-02-02 03:48郑希付
心理学报 2014年4期
孙 楠 郑希付
(1广州大学心理学系, 广州 510006) (2华南师范大学心理学院, 广州 510631)
1 引言
条件性关联学习模型以典型的巴普洛夫条件反射为基础, 主要研究记忆的习得、存储、提取和消退等过程。该模型被认为(Kim & Richardson,2009; Kim & Jung, 2006; LeDoux, 2000; Maren,2001)是研究创伤后应激障碍的典型动物模型,一般包括两个阶段, 关联学习阶段(或者称为习得阶段)和消退阶段。在关联学习阶段, 先对动物进行不可逃避非条件刺激(unconditioned stimulus, US,如电击)与条件刺激(conditioned stimulus, CS, 如灯光、声音或训练环境)的关联学习(CS-US)训练, 训练后当动物再次接触这个中性刺激 CS 时就会表现出条件性恐惧反应。消退阶段, 条件刺激 CS反复单独呈现而不匹配非条件刺激时(CS-no US), 那么动物先前习得的对 CS条件性恐惧反应会渐渐消退(Davis, Walker, & Myers, 2003)。动物对CS产生的各种生理反应(conditioned reaction, CR)被称为条件反应。早在1979年Bouton和Bolles两位学者已经发现, 消退过程不是自然发生的, 需要被试主动抑制之前获得的条件性关联记忆(CS-US), 继而形成一种新的CS-no US的消退记忆(extinction memory)(Bouton, García-Gutiérrez, Zilski, & Moody, 2006)。
随着条件性关联学习模型在人类的应用, 研究者开始使用各种视觉和听觉刺激作为 US, 代替既往动物研究中的电击刺激。人类的模型研究(Shanks& John, 1994; Wong, Bernat, Bunce, & Shevrin,1997)结果显示, 联想学习的成功取决与 CS-US之间的间隔, 间隔越短则联想学习越容易成功, 在CS和US间隔小于3秒的情况下, CS-US连续配对出现4~6次以上, 人类可以成功建立两者的条件性关联。刺激类型的不断扩展, 使得该模型在事件相关脑电技术的研究成为可能。研究者认为认知神经科学中事件相关电位技术(event-related potential,ERP)对关联学习的研究优于既往行为实验(Esteves,Dimberg, & Ohman, 1994, Esteves, Parra, Dimberg, &Ohman 1994; Ohman & Soares, 1993, 1994; Wong,Shevrin, & Williams, 1994)。Wong等研究者从1994年开始将条件性关联学习模型应用于人类认知神经的研究, 分别在1994年、1997年和2004年运用此模型进行了3个相关的ERP实验, 对条件性关联学习的早期神经机制进行研究。研究者主要分析的成分包括 P2, N2, P3和晚期正电位(late positive potential, LPP)等成分。其中P2和N2的波幅和潜伏期主要反应条件性关联学习中晚期的神经传导情况。P3和LPP成分则主要反应晚期的学习和消退的程度。具体到各个成分与条件性关联学习和消退过程的对应, 由于研究者使用的刺激类型不同,得到的结果和给出的解释也不尽相同。
之前有大量研究表明, 男性和女性在完成某些学习任务时存在一定差别(Spence & Spence, 1966;Stark et al., 2006)。Dalla和Shors在2009年对经典和操作性条件作用的学习过程中的性别差异进行研究, 结果发现:女性对操作性条件反射的关联学习比男性快, 保持的时间也比男性更久, 然而在消退训练中, 女性则需要更长的时间才能取得明显的消退效果。动物研究中(Barker & Galea, 2010; Beck,Brennan, & Servatius, 2002), 雄鼠对针刺的条件性关联反应较雌鼠更强烈, 而消退阶段较雌鼠更早消退。以上大量研究多集中在对恐惧记忆消退的性别差异方面, 而生活中, 消退异常主要包括两类, 记忆消退异常(如创伤后应激障碍)和行为消退异常(如特定情景下强迫行为的出现)。两类外在表现不同的消退异常无法分割对待, 行为消退异常通常伴有记忆关联的异常, 强迫行为的研究要以记忆消退异常为基础。本研究在记忆消退的性别差异研究基础上, 旨在探讨特定背景下的关联行为和消退的性别差异。根据临床经验, 强迫行为通常是在一定情绪背景或者特定情景下出现的, 考虑到这一现实情况, 本研究使用负性图片作为条件背景, 使背景和声音之间产生条件性关联, 被试听到声音后做出按键这一条件反应。
前人运用条件性关联模型对男性和女性在恐惧记忆消退方面做了大量研究, 这为本研究提供了理论基础。我们假设, 在负性条件背景下, 女性比男性对一般刺激建立的条件反应更难消退, 可能在消退阶段较男性出现更大的P3和LPP成分波幅。将条件性关联学习模型和事件相关电位技术结合探究其神经活动的性别差异, 是本研究的创新之处。参照既往研究(Albert, López-Martín, & Carretié,2010), 使用负性图片作为 CS, 同时为了保证负性图片对被试有稳定的影响, 在CS后立即出现US。采用Wong等研究者在2004年实验中的的条件性关联学习模型, 分成3个阶段(习惯阶段, 关联学习阶段, 消退阶段)进行研究, 同时参考 Neumann 和Kitlertsirivatana (2010)实验中对消退过程的控制。
2 材料和方法
2.1 被试和材料
共31个被试(15男, 16女), 均为在校大学生。平均年龄21±2.1岁。所有被试右力手。体重指数在18~24 kg/m之间。视力或校正视力正常, 无听力障碍。经检验, 以上两组年龄(t
(29) = 0.59), 体重指数(t
(29) = 1.25)均无显著性(p
>0.05)。CS为负性图片3张, 取自国际图片库, 和文献(Albert et al., 2010)中的图片相同, 并进行了重新评定。3张负性图片效价[F
(2, 87) = 0.003,p
= 0.99]和唤醒度[F
(2, 87) = 0.002,p
= 0.99]的方差分析结果均不显著, 保证同一属性的 CS能够诱发稳定的情绪和唤醒。US为中性声音, 实验中呈现声压强 85 dB, 频率为1000 Hz的中性声音。2.2 程序
实验程序用 E-prime编制, 程序分为 3个阶段。第一阶段(习惯阶段/部分实验称为暴露阶段),注释点1000~1400 ms的随机, 紧跟一个300 ms的负性图片(CS), 让被试自然观看, 3张图片分别连续出现13次, 13次和14次, 共40个trails, 顺序做被试间平衡。第二个阶段(关联学习阶段/部分实验称习得阶段)和第一阶段不同的是每张图片呈现结束, 都会出现一个1000 Hz的中性声音作为US,持续40 ms, US出现40次, 被试的任务是听到声音又快又准的按键, 按键后, 进入下一个trail。在正式实验前, 先进行练习。要求被试, 每张图片呈现结束都会跟随一个中性声音刺激, 虽然每张图片后都会跟随声音, 但是不能抢按键, 要认真自然观看图片, 被试的正确率达到 99%后进入正式实验。
这一阶段结束后, 参照既往研究(Harris, 2005),进行一个行为测验。这个行为测验是为了测试被试是否成功建立了 CS-US的关联学习。不能通过测试者数据被剔除。被试完成以下问题, (1)请说出中性声音什么时候出现; (2)你如何得到问题1的答案;(3)此阶段的实验, 哪个信号出现后, 会出现中性声音; (4)负性图片和中性声音之间的关系如何?这 4个问题通过后, 进入第三阶段实验。第三阶段(消退阶段), 告知被试声音可能出现也可能不出现, 让被试注意观看图片的同时, 注意声音, 听到声音就又快又准按键, 没有声音则不按键。但是声音没有出现, Tail数为40。根据Neumann和Kitlertsirivatana(2010)的研究, 为了保证消退训练的效果, 3张图片各连续5次呈现, 顺序做被试间平衡, 20个tail后的中间休息要向被试提问, 是否听到中性声音, 阶段结束要汇报中性声音的个数。关联学习阶段流程如图1所示。
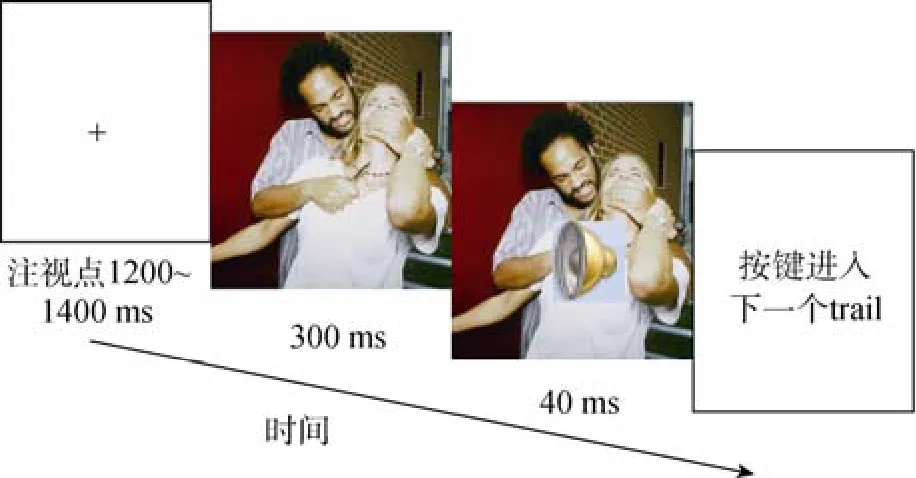
图1 关联学习阶段流程图
2.3 脑电记录
采用 BP ERP记录与分析系统, 按国际 10-20系统扩展的32导电极帽记录EEG。脑电记录时各电极参考于左侧乳突, 离线分析时通过对TP8的再参考, 实现以TP8和左侧乳突的平均值为参考。前额中央接地。垂直眼电(VEOG)电极分别安放在左眼的上下1.5 cm处。水平眼电(HEOG)电极分别置于左右眼外眦。所有电极阻抗降到5 kΩ。记录采用的滤波宽带为0.01~250 Hz。脑电与眼电均采用500 Hz采样频率连续采样。离线分析的数字滤波为低通16 Hz (24dB /octave)。波幅大于±100 μV的trial被视为伪迹剔除。
2.4 数据分析
行为数据采用E-prime软件采集, 用SPSS统计软件对关联学习阶段的反应时进行t
检验。脑电分析采用任务锁定, 三阶段的数据分析均从CS出现的-100 ms开始至出现700 ms结束。参照既往的研究报告(Wong, Bernat, Snodgrass, &Shevrin, 2004)本研究主要分析P2, N2, P3和LPP四个成分进行分阶段数据分析, 主要分析中线附件的9 个电极点: F4、F3、C3、C4、P3、P4、Fz、Cz和Pz。分别对关联学习阶段和消退阶段的P2、N2、P3、LPP四种成分一定时间窗内的波幅和潜伏期进行 2(性别) × 3(左右位置:左、中、右) × 3(前后位置:F区域、C区域、P区域)的重复测量方差分析, 并对p
值进行Green-house-Geisser矫正。为了检验消退效果同时排除习惯化效应, 对以上 9个电极P3/LPP波幅平均进行2阶段(习惯阶段、消退阶段)× 2性别(男、女)的重复测量方差分析, 并对p
值进行Green-house-Geisser矫正。3 结果
3.1 行为数据
无论男性和女性, 在关联学习阶段对 US按键正确率均在 100%, 反应时均小于 550 ms, 具体行为数据见表1组间t
检验显示男性和女性对US按键的反应时差异不显著,t
(29) = -0.36,p
= 0.72。
表1 男性和女性对US的反应时(M±SD)和错误率
3.2 ERP结果
ERP基本特征
根据当前研究的ERP总平均图, 在关联学习和消退阶段均观察到 P2、N2、P3和LPP (见图2、图3、图4), P2波幅在关联学习和消退阶段均是在 140~230 ms时间窗差异最大; 关联学习阶段, 阶段 N2 波幅在 300~360 ms时间窗差异最大, P3波幅在370~430 ms时间窗差异最大;消退阶段, N2在 230~320 ms时间窗差异最大;P3/LPP在350 ms时间窗后差异最大。这与Wong等(2004)研究者联想学习相关的ERP研究所观察的波形以及时间窗基本一致。关联学习阶段
N2
对 US按键任务诱发的 N2波幅在 300~360 ms时间窗的平均波幅进行重复测量方差分析得出, N2性别主效应显著,F
(1,29) = 3.59,p
= 0.03,其中男:-11.08±8.73 μV; 女:-7.86±5.37 μV。左右位置的主效应显著,F
(2,58) = 3.26,p
= 0.07, 最大波幅出现在中线区域(-11.68±5.88 μV), 而左侧(-7.89±6.41 μV)区域和右侧区域(-8.68±4.85 μV)间无显著差异。前后电极点主效应不显著,F
(2,58) =0.32,p
= 0.58。性别和前后位置的交互作用不显著:F
(2,58) = 0.67,p
= 0.43。性别和左右位置的交互作用不显著,F
(2, 58) = 0. 59,p
= 0.53。左右位置和前后位置的交互作用显著,F
(4,116 ) = 7.22,p
<0.001,最大波幅出现在 Fz 点(-10.45±5.65 μV)。三因素间的交互作用不显著,F
(4, 116) = 0.08,p
= 0.78。将300~360 ms时间窗内进行N2波峰标记, 将男性和女性N2的潜伏期进行两样本t
检验, 结果显示男性的潜伏期(M
= 0.32,SD
= 0.46)和女性的潜伏期(M
=0.32,SD
= 0.28), 差异不显著t
(29) = -0.04,p =
0.97。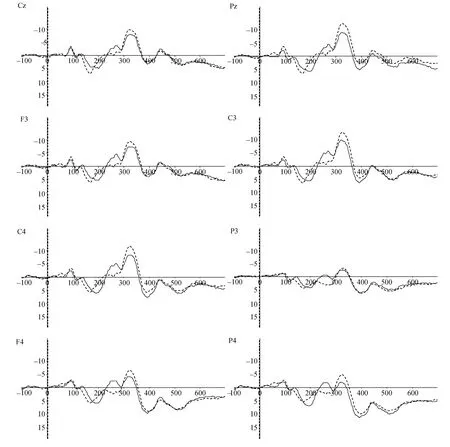
图2 男性(虚线)和女性(实线)在习得阶段ERP总平均图
P2
对US按键任务诱发的P2波幅在140~230 ms时间窗的平均波幅进行重复测量方差分析得出,P2性别主效应不显著,F
(1,29) = 1.94,p
= 0.17, 左右位置主效应不显著,F
(2,58) = 0.71,p
= 0.50, 前后位置主效应不显著,F
(2, 58) = 1.98,p
= 0.82, 性别和左右位置交互作用显著(F
(2, 58) = 4.57,p
=0.04), 男女在右侧脑区差异显著(p
<0.05), 而在中线和左侧脑区差异不显著(p
>0.05), 性别和前后位置(F
(2, 58) = 0.55,p
= 0.47)的交互作用不显著。将140~230 ms时间窗内进行P2波峰标记, 将男性和女性P2的潜伏期进行两样本t
检验, 结果显示男性的潜伏期(M
= 0.17,SD
= 0.15)比女性的潜伏期(M
= 0.22,SD
= 0.18)短, 并且差异显著t
(29) = -0.78,p
= 0.44。P3
对370~430 ms时间窗的P3平均波幅分析结果显示, 性别主效应显著,F
(1,29) = 55.10,p
<0.001, 其中女性(5.33±2.49 μV)显著高于男性(2.52±1.09 μV)。左右位置主效应显著,F
(2,58) = 5.07,p
= 0.01, 其中右侧位置(4.84±2.90 μV)明显大于左侧位置(3.08±1.78μV)波幅和中线位置(3.97± 2.17 μV),前后位置的主效应不显著,F
(2, 58) = 0.005,p
= 0.95,性别和前后位置的交互作用不显著:F
(2, 58) = 0.008,p
= 0. 93。性别和左右位置的交互作用不显著,F
(2,58) = 0.50,p
= 0. 61。左右位置和前后位置的交互作用显著,F
(4, 116 ) = 4.23,p
= 0.049。三因素间的交互作用不显著,F
(4, 116) = 1.76,p
= 1.19。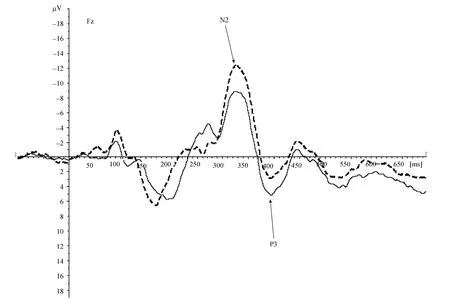
图3 习得阶段, 男性(虚线)和女性(实线)的ERP总平均波幅在Fz点的差异
消退阶段
P2
在140~230 ms时间窗的平均波幅进行重复测量方差分析得出, P2性别主效应显著F
(1,29) =16.19,p
<0.01, 男性(5.93±3.05 μV)波幅明显大于女性(4.37±2.41 μV), 左右位置(F
(2,58) = 0.23,p
=0.64)和前后位置(F
(2,58) = 0.71,p
= 0.41)的主效应不显著, 性别和左右位置(F
(2,58) = 0.001,p
= 0.97)前后位置(F
(2,58) = 1.99,p
= 0.17)交互作用均不显著, 性别、左右和前后位置三者的交互作用不显著(F
(4,116) = 2.34,p
= 0.11)。将140~230 ms时间窗内进行P2波峰标记, 将男性和女性P2的潜伏期进行两样本t
检验, 结果显示男性的潜伏期(M
= 0.17,SD
= 1.10)和女性的潜伏期(M
= 0.19,SD
= 0.42)差异不显著t
(29) = -0.05,p =
0.97。N2
N2波幅在230~320 ms时间窗的平均波幅进行方差分析得出, N2性别主效应不显著,F
(1,29)= 0.04,p
= 0.95; 左右位置主效应不显著,F
(2,58) =1.06,p
= 0.36; 前后位置主效应显著,F
(2,58) = 4.30,p
= 0.04。 其中F区(-3.08±2.18 μV)、C区(-3.02±2.03 μV)的波幅大于 P (-2.15±1.76 μV)区的波幅, 性别和左右位置(F
(2,58) = 0.03,p
= 0.87)前后位置交互作用均不显著(F
(2,58) = 0.09,p
= 0.91), 三者的交互作用不显著,F
(4,116) = 0.41,p
= 0.66。将230~320 ms时间窗内进行N2波峰标记, 男性和女性N2的潜伏期进行两样本t
检验, 结果显示女性的潜伏期(M
= 0.27,SD
= 0.02)和男性的潜伏期(M
=0.34,SD
= 0.12), 差异显著,t
(29) = 2.18,p
= 0.04。P3/LPP
350 ms~700 ms时间窗的平均波幅方差分析得出, 性别主效应显著,F
(1, 29) = 42.56,p
<0.01。左右位置主效应显著,F
(2,58) = 4.10,p
=0.02, 其 中 中 线 位 置 (4.18±2.26 μV)大 于 左 侧(3.25±2.00 μV)和右侧(3.09±1.79 μV)。前后位置主效应不显著,F
(2,58) = 1.52,p
= 0.23。性别和左右位置(F
(2,58) = 0.23,p
= 0.64)和前后位置(F
(2,58) =0.05,p
= 0.94)的交互作用均不显著。三者的交互作用不显著,F
(4,116) = 1.85,p
= 0.17。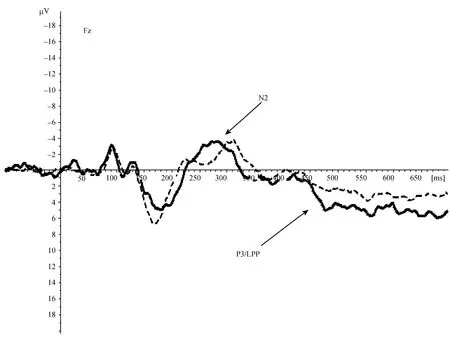
图4 消退阶段男性(虚线)和女性(实线)在FZ点ERP总平均图
P3/LPP波幅的2阶段(习惯阶段、消退阶段) × 2性别(男、女)的方差分析结果, 性别主效应显著,F
(1,29) = 6.01,p
= 0.02, 阶段反应主效应显著,F
(1,29) = 6.49,p
= 0.01。两者交互作用显著,F
(1,29)= 5.56,p
= 0.02, 其中男女性在习惯阶段的差异不显著(p
>0.05), 而在消退阶段的差异显著(p
<0.05);女性在习惯和消退阶段差异显著(p
<0.05), 女性在消退阶段的波幅(4.58±1.88 μV)明显大于在习惯阶段的波幅(2.31±1.35 μV), 男性在两阶段的差异不显著(p
>0.05)。4 讨论
本研究目的, 考察男女性在负性背景下条件性关联学习和消退过程的神经活动差异。研究发现,在关联学习阶段, 男性存在更短的 P2潜伏期和更大的 N2波幅, 这暗示男性可能较女性更快的探测到条件刺激(CS)和非条件刺激(US)之间的联系, 而女性出现较男性更大的 P3波幅, 暗示女性在晚期才探测到条件刺激和非条件刺激之间的联系, 进一步表明女性可能慢于男性建立这种关联学习。消退过程中, 已经建立的CS-US关联消失, 形成一种新的 CS-no US的关联, 男性在早期对这一过程做出反应, 主要表现在男性较女性更大的 P2波幅, 而女性则在稍晚期对这一过程做出反应, 并且需要付出较男性更大的认知资源, 才能适应这一新的关联,主要表现在女性较男性更大的 P3/LPP的波幅。与之前研究相比, 本研究通过对关联学习和消退神经活动的研究, 得到女性和男性在同一阶段的不同时间进程上表现不同, 这是行为实验无法达到的, 可以在一定程度上解释部分行为研究结果背后的潜在神经基础。并且在前人研究的基础上, 对ERP各个成分与关联学习和消退过程更好的对应做出一定贡献。
4.1 关联学习阶段的性别差异
关联学习阶段男性的 ERP中晚期成分活跃,表现为男性较女性出现更短的 P2潜伏期和更大的N2波幅。Wong等研究者在2004年的消退ERP研究中认为, P2反应条件刺激(CS)和非条件刺激(US)之间的关联程度。而Bernat在2001年的研究指出,P2可以反应关联学习的传导过程, 这一结论早在1968年Shevrin和Fritzler的研究中, 对P2有类似的解释。本研究中, US在CS出现的末期出现, 相当于以CS-US复合物的形式出现, 可以认为P2在一定程度上反应了条件刺激和非条件刺激之间的关联学习情况, 其中 P2潜伏期反应条件反应的速度, P2波幅反应条件性关联的程度。既往研究中N2成分被认为是注意与任务相关的信息加工过程、同时抑制无关信息, 体现反应冲突以及冲突监控过程中注意资源分配等(Barry, 2009; Zimmer & Demmel,2000; Bledowski, Prvulovic, Goebel, Zanella, &Linden, 2004), 本研究N2的结果可以认为, 与女性相比, 男性抑制负性背景信息诱发的冲突更大, 需要更大的认知资源。结合P2和N2的结果, 可以认为男性在关联学习的中晚期产生更大的反应, 体现了男性可能快于女性探测到 CS-US之间的关联,这可能是男性能够较女性更快形成关联学习的神经证据。
关联学习阶段 ERP的晚期成分表现为, 女性较男性出现更大的 P3波幅。Wong等 (2004)研究结果认为 P3的波幅在一定程度上反应了条件性关联学习的结果, 这与前人(Begleiter & Platz, 1969;Wong et al., 1994)研究结果一致。但是研究者没有明确说明 P3成分如何具体反应学习的结果, 比如反应的是关联学习的速度还是程度。而 Pauli和Röder (2008)研究认为, P3可以反应条件性关联学习过程中, 认知资源的分配, P3波幅越大则表明分配的认知资源越多。相对于男性在 ERP中晚期成分N2和P2上反应更大, 而女性则在ERP晚期成分P3反应更大, 我们认为此阶段男性和女性在P3上存在的差异很可能表示为, 在关联学习的晚期女性较男性投入更多的认知资源, 进一步证实女性可能慢于男性探索到这种CS和US之间的关联学习,并且保持时间更久。结合关联学习的早期和中晚期的 ERP各个成分的结果, 我们得到了以时间进程为线索的男性和女性神经活动的差别。
4.2 消退阶段的性别差异
既往研究(Bouton, 2002; Quirk, 2002; Rescorla,2004; Quirk, Garcia, & González- Lima, 2006)认为,消退阶段比之前的关联学习阶段更加复杂, 被试要切断之前的CS-US关联学习, 形成新的CS-no US的关联学习, 这一过程不是自然发生, 而是需要认知资源的。根据消退相关 ERP研究中(Driver,Vuilleumier, Eimer, & Rees, 2001; Pizzagalli, Greischar,& Davidson, 2003), 认为P1、P2等早中期成分体现消退阶段的神经传导过程。由于研究者采用的模型类似, 但采用的刺激类型各异, 对于消退的ERP早期成分的解释不尽相同。本研究中, 消退早期男性较女性表现出更大的 P2波幅。这一结果表明, 男性可能较女性更早的对消退过程做出反应, 或者说男性可能更早的察觉到新的关联形式的出现, 并在这一过程中释放了更多的认知资源。
为了证明被试成功的进行了关联学习, 我们对比了刺激呈现流程一致的习惯阶段和消退阶段的P3/LPP成分, 这一成分在以往的关联学习研究中被认为一定程度上反映了记忆的消退程度(Pizzagalli et al., 2003; Wong et al., 2004)。结果发现,P3/LPP的波幅有明显的阶段效应, 即消退阶段的晚正成分要大于习惯阶段, 这可以被理解为被试为了打破已建立的关联学习, 需要投入更多的心理资源, 同时也说明本研究设计的有效性, 基本排除了图片本身和习惯化引起的性别差异。如果因为习惯化引起的性别差异, 习惯化使负性图片引起的 P3波幅有逐渐减小的趋势, 消退阶段的波幅应该小于习惯阶段的波幅。另外, 男女性的 P3/LPP波幅在消退阶段差异显著而习惯阶段不显著, 这从另一方面证明存在成功的关联学习。女性较男性表现出更大的 P3/LPP波幅, 我们认为在消退的晚期, 女性为了完成新的关联学习, 同时之前的 CS-US关联学习尽快消退, 需要投入比男性更多的资源。这可能意味着负性背景对男性和女性的影响存在时间上的差异, 男性能够更好的克服负性背景的影响,在较早期对消退产生反应, 并投入比女性更多的资源, 女性则在晚期对消退产生反应, 在晚期甚至更晚期对消退任务投入更多的资源或者更多的努力。提示我们注意, 具有一定情绪基础的女性, 在进行消退学习中, 控制情绪对于女性可能较男性更加重要, 这对暴露治疗过程中, 男性和女性的差异化治疗提供了一定的理论基础。
5 结论
本研究的创新之处在于发现了负性背景下男性和女性在条件性关联学习和消退过程中神经活动的差异, 并且以时间进程为线索,得到男性和女性在同一阶段的不同时程表现出不同的神经活动:在关联学习和消退阶段, ERP中晚期成分N2和P2波幅和潜伏期的性别差异和晚期成分P3和LPP波幅的性别差异, 反应了男性和女性神经活动差异的复杂性。在前人研究的基础上, 综合本研究两阶段脑电数据, 我们认为负性背景下的关联学习和消退过程不是一个简单的过程, 不能单纯给出男性容易习得或者女性难消退这一简单的结论, 不同时间进程会出现不同的神经活动差异。ERP成分与关联学习和消退程度的严格对应关系仍然需要今后的研究继续探讨, 对于负性背景下, 男性和女性哪个群体更容易建立学习连接和消退可能需要更深一步的认知神经实验研究。
Albert, J., López-Martín, S., & Carretié, L. (2010). Emotional context modulates response inhibition: Neural and behavioral data.Neuroimage,49
(1), 914–921.Barker, J. M., & Galea, L. A. M. (2010). Males show stronger contextual fear conditioning than females after context pre-exposure.Physiology & Behavior,99
, 82–90.Barry, R. J. (2009). Evoked activity and EEG phase-resetting in the genesis of auditory Go/NoGo ERPs.Biological Psychology,80
, 292–299.Beck, K. D., Brennan, F. X., & Servatius, R. J. (2002). Effects of stress on nonassociative learning processes in male and female rats.Integrative Physiological and Behavioral Science,37
(2), 128–139.Begleiter, H., & Platz, A. (1969). Evoked potentials:Modifications by classical conditioning.Science,166
,769–771.Bledowski, C., Prvulovic, D., Goebel, R., Zanella, F. E., &Linden, D. E. J. (2004). Attentional systems in target and distractor processing: A combined ERP and fMRI study.NeuroImage,22
, 530–540.Bouton, M. E. (2002). Context, ambiguity, and unlearning:Sources of relapse after behavioral extinction.Biological Psychiatry,52,
976–986.Bouton, M. E., García-Gutiérrez, A., Zilski, J., & Moody, E. W.(2006). Extinction in multiple contexts does not necessarily make extinction less vulnerable to relapse.Behaviour Research and Therapy, 44
, 983–994.Davis, M., Walker, D. L., & Myers, K. M. (2003). Role of the amygdala in fear extinction measured with potentiated startle.Annals of the New York Academy Science,985
,218–232.Driver, J., Vuilleumier, P., Eimer, M., & Rees, G. (2001).Functional magnetic resonance imaging and evoked potential correlates of conscious and unconscious vision in parietal extinction patients.NeuroImage,14
, S68–S75.Esteves, F., Dimberg, U., & Öhman, A. (1994). Automatically elicited fear: Conditioned skin conductance responses to masked facial expressions.Cognition and Emotion,8
, 393–413.Esteves, F., Parra, C., Dimberg, U., & Öhman, A. (1994).Nonconscious associative learning: Pavlovian conditioning of skin conductance responses to masked fear-relevant facial stimuli.Psychophysiology,31
, 375–385.Harris, J. B. (2005). Differential conditioning of alpha amplitude: A fresh look at an old phenomenon.Clinical Neurophysiology,116
, 1433–1443.Kim, J. H., & Richardson, R. (2009). New findings on extinction of conditioned fear early in development:Theoretical and clinical implications.Biological Psychiatry,67
, 297–303.Kim, J. J., & Jung, M. W. (2006). Neural circuits and mechanisms involved in Pavlovian fear conditioning: A critical review.Neuroscience and Biobehavioral Reviews,30
, 188–202.LeDoux, J. E. (2000). Emotion circuits in the brain.Annual Review of Neuroscience,23
, 155–184.Maren, S. (2001). Neurobiology of Pavlovian fear conditioning.Annual Review of Neuroscience,24
, 897-931.Neumann, D. L., & Kitlertsirivatana, E. (2010). Exposure to a novel context after extinction causes a renewal of extinguished conditioned responses: Implications for the treatment of fear.Behaviour Research and Therapy,48
,565−570.Ohman, A., & Soares, J. J. F. (1993). On the automatic nature of phobic fear: conditioned electrodermal responses to masked fear-relevant stimuli.Journal of Abnormal Psychology,102
, 121–132.Ohman, A., & Soares, J. J. F. (1994). Unconscious anxiety:Phobic responses to masked stimuli.Journal of Abnormal Psychology,
103, 231–240.Pauli, W. M., & Röder, B. (2008). Emotional salience changes the focus of spatial attention.Brain Research,1214
,94–104.Pizzagalli, D. A., Greischar, L. L., & Davidson, R. G. (2003).Spatio-temporal dynamics of brain mechanisms in aversive classical conditioning: High-density event-related potential and brain electrical tomography analyses.Neuropsychologia,41
, 184–194.Quirk, G. J. (2002). Memory for extinction of conditioned fear is longlasting and persists following spontaneous recovery.Learning and Memory, 9
, 402–407.Quirk, G. J., Garcia, R., & González-Lima, F. (2006).Prefrontal mechanisms in extinction of conditioned fear.Biological Psychiatry, 60
, 337-343.Rescorla, R. A. (2004). Spontaneous recovery.Learning and Memory,11
, 501–509.Shanks, D. R., & St John, M. F. (1994). Characteristics of dissociable human learning systems.Behavioral and Brain Sciences, 17
, 367–395.Shevrin, H., & Fritzler, D. E. (1968). Visual evoked response correlates of unconscious mental processes.Science,161
,295–298.Spence, K. W., & Spence, J. T. (1966). Sex and anxiety differences in eyelid conditioning.Psychological Bulletin,65
, 137−142.Stark, R., Wolf, O. T., Tabbert, K., Kagerer, S., Zimmermann,M., Kirsch, P.,... Vaitl, D. (2006). Influence of the stress hormone cortisol on fear conditioning in humans: Evidence for sex differences in the response of the prefrontal cortex.Neuroimage, 32
, 1290−1298.Wong, P. S., Bernat, E., Bunce, S., & Shevrin, H. (1997).Brain indices of nonconscious associative learning.Consciousness and Cognition,6
, 519–544.Wong, P. S., Bernat, E., Snodgrass, M., & Shevrin, H. (2004).Event-related brain correlates of associative learning without awareness.International Journal of Psychophysiology,5
3, 217–231.Wong, P. S., Shevrin, H., & Williams, W. J. (1994). Conscious and nonconscious processes: an ERP index of an anticipatory response in a conditioning paradigm using visually masked stimuli.Psychophysiology,31
, 87–101.Zimmer, H., & Demmel, R. (2000). Habituation and laterality of orienting processes as reflected by slow negative waves.Biological Psychology,53
, 161–176.猜你喜欢
健康体检与管理(2022年4期)2022-05-13
中国药学药品知识仓库(2021年18期)2021-02-28
初中生学习指导·中考版(2020年2期)2020-09-10
初中生世界·八年级(2019年8期)2019-08-29
价值工程(2018年25期)2018-09-26
学苑创造·A版(2018年11期)2018-02-01
数学大王·低年级(2017年12期)2017-12-26
中国现代医生(2017年24期)2017-10-17
读者(2017年5期)2017-02-15
科技视界(2016年6期)2016-07-12
