
北极新奥尔松Austre Lovénbreen冰川退缩迹地不同演替阶段的植物组成与植被群落特征分析
2014-03-08 01:25姚轶锋曹叔楠彭方李承森
极地研究 2014年3期
姚轶锋 曹叔楠 彭方 李承森
(1中国科学院植物研究所系统与进化植物学国家重点实验室,北京100093;2中国科学院微生物研究所真菌学国家重点实验室,北京100101;3武汉大学生命科学学院,湖北武汉430072)
0 引言
北极是地球上的气候敏感地区,也是研究全球气候变化的关键地区。北极所处地理位置特殊,特有的生态、气候和自然环境变化,在全球变化,尤其是在全球及区域气候变化中有重要作用[1]。北极的气候和环境变化,将影响着全球气候和环境变化,并将直接影响着人类的生存和未来。这种变化也会影响中国的气候、环境和可持续发展。
冰川退缩和冻土层消融等证据表明北极地区正在经历快速的变暖过程[2-4],其结果将显著地影响到北极生态系统中植被的组成、分布和发展变化[5-9]。北极的环境条件恶劣(例如:极端寒冷、半年白昼和黑夜交替),植物的生长季较短,获得的有效养分有限,因而植物的生长率相对低下,植被的演替速率缓慢[10-11]。然而,全球变暖的结果将加速植物的生长和植被的演替速率[12]。因此,在北极布设观测样方对植被群落变化进行长期监测,可以在一定程度上反映全球气候变化的趋势。
冰川退缩后裸露的陆地为地衣和植物的生长提供了新的栖息地。根据冰川退缩的年代可以为植被演替提供准确的时间序列,因此,冰川退缩后的裸地也是研究植被发生、发展的理想区域。前人对北极新奥松地区Midre Lovénbreen冰川(简称M冰川)和Austre Lovénbreen冰川(简称A冰川)前沿裸地上不同区域环境的植被组成进行过对比研究[13]。本文主要对A冰川退缩迹地上不同演替阶段的植物组成与植被群落特征进行调查,为今后植被的长期监测提供基本数据。
1 研究区域概况
斯瓦尔巴群岛(74°N—81°N,10°E—35°E)处于巴伦支海和格陵兰海之间,由斯匹次卑尔根岛、东北地岛、埃季岛等三个大岛和数十个小岛组成,总面积约6.2×104km2,其中60%被冰川所覆盖。新奥尔松(78°55′N,11°56′E)位于斯瓦尔巴群岛的西部(图1a),是北极的科考基地。该地区具有极地-海洋性气候特征,由于受北大西洋暖流的影响,气温要比北极同纬度其他地区温和许多。最冷月均温(2月)-14℃,最热月均温(7月)5℃。年均温(2001—2008年)-4.2℃,年均降水量 433 mm,每年7月初冰雪开始融化,9月中旬开始积雪[14]。新奥尔松地区的植物种类相对较少,主要以北部极地苔原植物区系和极地沙漠植物区系为主,该地区常见的苔原植物包括杨柳科极柳(Salix polaris)、虎耳草科挪威虎耳草(Saxifraga oppositifolia)和零余虎耳草(Saxifraga cernua)、石竹科北极鼠耳草(Cerastium arcticum)和无茎蝇子草(Silene acaulis)、蔷薇科仙女木(Dryas octopetala)、罂粟科斯瓦尔巴罂粟(Papaver dahlianum)以及莎草科苔草(Carexmisandra)等。
2012年植被调查的区域位于A冰川前沿,该冰川与北极黄河站的直线距离为6.2 km,为多热型山谷冰川,周围山峰的最高海拔为880 m,冰川最高海拔为600 m。冰川表面均较为平整,有少量碎屑物质,且均发育少量冰川裂隙[15]。A冰川不同年代的冰缘线位置见图1b。
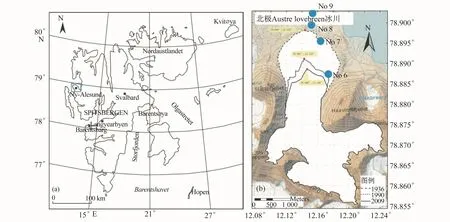
图1 北极新奥尔松地理位置和样方位置示意图Fig.1.Map showing the position of Ny-Ålesund and quadrats
2 材料与方法
从A冰川的地形图上(图1b)可以明显地反映出冰川退缩的趋势。2009年冰缘线位置根据中国北极科考队员在A冰川表面野外采集的实测GPS数据确定。1936年和1990年冰缘线位置引自挪威极地研究所1990年和2008年分别出版的北极Svalbard地区A7(Kongsfjorden)片区地形图,这两幅地形图都是基于航片,利用摄影测量法汇编的地形图[16-17]。斯瓦尔巴地区小冰期结束以后,A冰川逐渐衰退,通过对比Moreau等2009年论文中的图1(小冰期结束以来M冰川和A冰川退缩的不同阶段和位置)[13],初步确定8号样方可能位于小冰期结束时的冰缘线附近。但目前国际上对该地区小冰期结束时间至今存在争议,有学者认为结束偏早(1880年 左 右)[18],也 有 学 者 主 张 结 束 稍 晚(1918年[19]、约 1900—1920年[20])。鉴于小冰期结束时间至今没有定论,所以本文暂用可能在小冰期结束时冰缘线附近来描述8号样方的位置(可能约90—130年前)。
在2012年7月北极黄河站现场考察中,我们采用样线法沿A冰川前沿至海岸横断面建立了4个样方。样方分别位于1936年和1990年冰缘线、小冰期结束时的冰缘线附近,以及退缩时间可能大于90—130年的位置(表1,图 1b)。样方总大小为1 m×1.5 m,其中包括1个1 m×1 m的小样方,用于开展苔原植物生长和植被演替对冰川退缩响应的研究;两个0.5 m×0.5 m的小样方,供其他学科取样开展土壤微生物和重金属等方面的研究。为了方便统计和定位样方内每种植物和地衣,将1 m×1 m样方细分为100个10 cm×10 cm小格。详细调查每个样方内维管植物种类和个体数,计算其多度,同时调查样方内的地衣种类及其盖度。测定每个样方地表以下5 cm处土壤的温度、湿度和pH值。

表1 样方位置信息表Table 1.Location information of the quadrats
3 结果
3.1 物种组成及其差异性
4个样方共统计维管植物2 041株(丛),鉴定出9科14种。其中木本植物2种:极柳、仙女木,草本植物12种:虎耳草科3种(挪威虎耳草、零余虎耳草、丛生虎耳草)、石竹科3种(米努草、漆姑草、无茎蝇子草)、蓼科2种(珠芽蓼、冰岛蓼)、十字花科、玄参科、莎草科和禾本科各1种。共鉴定地衣7种,包括5种壳状地衣:寒生肉疣衣、藓生茶渍衣、鸡皮衣、脐鳞衣、橙衣,2种叶状地衣:雪黄岛衣、刺岛衣。另外有黑色壳状地衣未定种1种。样方内常见维管植物和地衣见图2。
冰川退缩迹地演替系列上,物种组成表现出明显差异性(图3),随演替时间增加,物种丰富度增加。代表冰川迹地形成时间22年时(6号样方),物种组成单一。7—9号样方分别代表迹地形成时间长达75年、可能约90—130年前、可能大于90—130年前,与6号样方相比,样方内物种组成更加丰富(5—8种),而且物种更替明显。由7号样方以极柳和黄葶苈为主,到8、9号样方以极柳占绝对优势。相应地,地衣也由壳状地衣为主,过渡到叶状地衣逐渐出现。从图3中可以看出,挪威虎耳草贯穿于迹地形成和植被演替的整个过程,但其优势地位逐渐被极柳取代。苔草、仙女木等物种通常出现在植被演替的成熟阶段。
3.2 群落结构特征及其差异性
冰川退缩迹地演替系列上,在植物群落结构方面也呈现出一定差异性。各样方的植物群落结构具有以下特征:
(1)冰川退缩22年后植被演替的初始阶段(6号样方),样方内碎石多,平均直径2—3 cm。样方内出现的植物种类少,只有先锋植物挪威虎耳草;另外有少量苔藓,未观察到地衣,但在样方周围有壳状地衣出现。
(2)冰川退缩75年后植被发育的情况(7号样方),植物种类和个体数明显比6号样方多,共出现7种植物,以木本植物极柳(35.5%)和草本植物黄葶苈(39.4%)占优势(图3)。出现壳状地衣如寒生肉疣衣和鸡皮衣等,并且地衣的盖度达到1.65%(图3)。
(3)冰川退缩可能约90—130年后植被发育的情况(8号样方),样方内的植物种类以极柳占绝对优势(94.2%)。各种地衣的盖度比7号样方均有所增加,出现叶状地衣——刺岛衣,但盖度较低(图3)。

图2 样方中常见维管植物和地衣种类.1—无茎蝇子草,2—挪威虎耳草,3—仙女木,4—米努草,5—漆姑草,6—极柳,7—苔草,8—珠芽蓼,9—黄葶苈,10—雪黄岛衣,11—藓生茶渍衣,12—寒生肉疣衣,13—刺岛衣,14—鸡皮衣,15—橙衣Fig.2.Common vascular plants and lichens in the quadrats.1—Silene acaulis,2—Saxifraga oppositifolia,3—Dryas octopetala,4—Minuartia rubella,5—Sagina nivalis,6—Salix polaris,7—Carexmisandra,8—Polygonum viviparum,9—Draba bellii,10—Flavocetraria nivalis,11—Lecanora epibryon,12—Ochrolechia frigida,13—Cetraria aculeate,14—Pertusaria sp.,15—Caloplaca sp.
(4)冰川退缩时间更长(可能大于90—130年)植被发育更好,从株(丛)数和多度来看,样方内植物以极柳占优势(79.1%);从盖度来讲,以仙女木占优势(约占样方的25%)。地衣的总盖度约占6.5%,其中寒生肉疣衣的盖度相比其他样方有所增加,黑色壳状地衣和鸡皮衣的盖度有所降低,出现叶状地衣——雪黄岛衣,其盖度较低(图3)。
3.3 土壤基本特性
冰川迹地形成22年时,样方内基质以粒径为2—3 cm的碎石为主,其风化程度和土壤发育程度较低,在碎石下层有少量粗骨性土壤。而随着冰川迹地形成时间长,演替时间增加,样方内基质的土壤化程度较高,尤其形成时间可能超过90—130年后(9号样方),基质演替为壤质土壤。短期调查初步结果(表2)表明,冰川迹地形成各时间序列上土壤的温度在7月未表现出差异,但湿度与pH值有差异,其中随迹地形成时间增加,pH值呈减小趋势,但总体表现为中性或弱酸性。

图3 冰川退缩迹地不同演替阶段的维管植物和地衣组成.维管植物种类:1—苔草,2—黄葶苈,3—仙女木,4—冰岛蓼,5—米努草,6—长毛马先蒿,7—北极早熟禾,8—珠芽蓼,9—漆姑草,10—极柳,11—零余虎耳草,12—丛生虎耳草,13—挪威虎耳草,14—无茎蝇子草.地衣种类:1—盖度合计,2—壳状地衣(未定种),3—寒生肉疣衣,4—鸡皮衣,5—藓生茶渍衣,6—橙衣,7—脐鳞衣,8—雪黄岛衣,9—刺岛衣Fig.3.Vascular plants and lichens during different succession stages on glacier foreland.Vascular plants:1—Carexmisandra,2—Draba bellii,3—Dryas octopetala,4—Koenigia islandica,5—Minuartia rubella,6—Pedicularis hirsuta,7—Poa arctica,8—Polygonum viviparum,9—Sagina nivalis,10—Salix polaris,11—Saxifraga cernua,12—S.cespitosa,13—S.oppositifolia,14—Silene acaulis.Lichens:1—Total coverage,2—Crustose lichen(unidentified species),3—Ochrolechia frigida,4—Pertusaria sp.,5—Lecanora epibryon,6—Caloplaca sp.,7—Rhizoplaca sp.,8—Flavocetraria nivalis,9—Cetraria aculeate

表2 样方内土壤温度、湿度和pH值测定结果Table 2.Results of soil temperature,humidity and pH value in the quadrats
4 讨论
前人对A冰川退缩迹地上植被的研究较少,仅见法国学者Moreau等有过报道。他们于1975年和2006年分别调查了M冰川、A冰川前沿至海岸横断面不同地形地貌单元下(如:冰碛带、冰水冲积平原、海岸阶地等)植物的组成及其变化,同时结合对应分析和等级分类方法探讨了植被的演替和植被类型的空间变化。其研究结果揭示不同调查地点的水流、迹地形成年代、土壤状况等环境条件是影响植被演替和植被类型的主要因素[13]。
我们的研究主要从演替的时间序列考虑,采用样线法在A冰川迹地不同形成年代位置布设样方,进行不同演替阶段的植物组成和植被群落结构的调查。初步结果表明:冰川迹地形成22年(6号样方),植被处于演替的初始阶段,植物组成和群落结构单一,主要是先锋植物挪威虎耳草和少量苔藓植物,附近观察到壳状地衣;冰川迹地形成75年(7号样方),植被处于演替的中间阶段,植物种类显著增加(7种),群落结构更加复杂,以木本植物极柳(35.5%)和草本植物黄葶苈(39.4%)为主,其他草本植物还有挪威虎耳草、零余虎耳草、丛生虎耳草、北极早熟禾、漆姑草,出现寒生肉疣衣和鸡皮衣等壳状地衣;冰川迹地形成可能约90—130年(8号样方),植被处于演替的成熟阶段,生境偏湿,植物组成和群落结构以极柳占绝对优势(94.2%),出现了少量珠芽蓼、冰岛蓼两种新类群,刺岛衣等叶状地衣开始出现;海岸阶地上的植被(9号样方)生长年限缺乏准确的年龄,推测可能超过90—130年,代表了演替的成熟阶段,生境偏干,植物组成和群落结构仍以极柳为主(79.1%),同时出现了苔草、仙女木、长毛马先蒿等指示干旱环境的类群,地衣的物种多样性和盖度明显增加。
冰川迹地上植被的演替是一个缓慢过程,从初始发展到成熟稳定阶段至少需要上百年时间。在整个演替过程中,驱动冰川迹地上物种的更替和群落结构改变的因素较多,其中区域环境(如:小气候、地形等)与冰川迹地基质(如:土壤粒度、有机质含量等)是非常重要的两方面。本文中测定的土壤温度、湿度和pH值等基本参数可能不是影响植被演替与植物定居的关键因素,今后有待于对冰川迹地基质进行全面的调查。
致谢 作者衷心感谢国家海洋局极地考察办公室对本研究工作的支持,感谢中国极地研究中心闫明研究员对A冰川冰缘线年代提出的修改意见,感谢武汉大学中国南极测绘研究中心艾松涛博士提供A冰川地形图,感谢2012年度中国北极黄河站科考队队友的大力协助。本论文部分样品的信息和数据由中国极地研究中心和中国南北极数据中心建设的“极地标本资源共享平台(http://birds.chinare.org.cn/)”提供。
1 陆龙骅,卞林根,效存德,等.极地大气科学与全球变化研究进展.应用气象学报,2006,17(6):743—755.
2 Watson R T,Zinyowera M C,Moss R H.The Regional Impacts of Climate Change:An Assessmentof Vulnerability.Cambridge:Cambridge University Press,1998.
3 IPCC.Climate Change,The Scientific Basis,Intergovernmental Panel on Climate Change.Third Assessment Report.Impacts,Adaptation and Vulnerability.Cambridge:Cambridge University Press,2001.
4 ACIA.Impacts of aWarming Arctic:Arctic Climate Impact Assessment.Cambridge:Cambridge University Press,2004.
5 Karlsen SR,Elvebakk A.A method using indicator plants tomap local climatic variation in the Kangerlussuaq/Scoresby Sund area,east Greenland.Journal of Biogeography,2003,30(10):1469—1491.
6 Wahren CH A,Walker M D,Bret-Harte M S.Vegetation responses in Alaskan arctic tundra after8 yearsof a summerwarming and winter snowmanipulation experiment.Global Change Biology,2005,11(4):537—552.
7 Walker M D,Wahren CH A,Hollister R D,etal.Plant community responses to experimentalwarming across the tundra biome.Proceedings of the National Academy of Sciences of the United States of America,2006,103(5):1342—1346.
8 Alsos IG,Eidesen P B,Ehrich D,et al.Frequent long-distance plant colonization in the changing Arctic.Science,2007,316:1606—1609.
9 Meltofte H,Christensen TR,Elberling B.High-Arctic Ecosystem Dynamics in a Changing Climate:Ten Years ofMonitoring and Research at Zackenberg Research Station,Northeast Greenland.Amsterdam:Academic Press,2008.
10 Churchill E D,Hanson H C.The concept of climax in arctic and alpine vegetation.The Botanical Review,1958,24(2-3):127—191.
11 Webber P J.Spatial and temporal variation of the vegetation and its production,Barrow,Alaska//Tieszen L L.Vegetation and Production Ecology of an Alaskan Arctic Tundra.New York:Springer-Verlag,1978,29:37—112.
12 Kojima S.A two-year change ofarctic vegetation as observed in a permanent plotestablished in Ny-Ålesund,Svalbard.Polar Bioscience,2002,15:123—128.
13 Moreau M,Laffly D,Brossard T.Recent spatial development of Svalbard strandflat vegetation over a period of31 years.Polar Research,2009,28(3):364—375.
14 Nakatsubo T,FujiyoshiM,Yoshitake S,etal.Colonization of the polarwillow Salix polaris on the early stage of succession after glacier retreat in the High Arctic,Ny-Ålesund,Svalbard.Polar Research,2010,29(3):385—390.
15 徐明星,闫明,任贾文,等.北极Svalbard地区Austre Lovénbreen和Pedersenbreen冰川表面物质平衡和运动特征分析.极地研究,2010,22(1):10—22.
16 Norwegian Polar Institute.Geological Map of Svalbard,Scale 1∶100000.Sheet A7,Kongsfjorden.Norsk Polarinstitutt,Tromsø.1990.
17 Norwegian Polar Institute.Geological Map of Svalbard,Scale 1∶100000.Sheet A7,Kongsfjorden.Norsk Polarinstitutt,Tromsø.2008.
18 Lefauconnier B.Fluctuations récentes des glaciers dansle Kongsfjord,79°N Spitsberg.Inter-Nord,1990,19:449—453.
19 Lefauconnier B,Hagen JO.Glaciers and climate in Svalbard:statistical analysis and reconstruction of the Brøggerbreenmass balance for the last77 years.Annals of Glaciology,1990,14:148—152.
20 Moreau M,Mercier D,Laffly D,et al.Impacts of recent paraglacial dynamics on plant colonization:A case study on Midtre Lovénbreen foreland,Spitsbergen(79°N).Geomorphology,2008,95(1-2):48—60.
猜你喜欢
林业勘查设计(2022年1期)2022-02-15
兰台内外(2021年33期)2021-12-17
乡村科技(2021年17期)2021-10-20
安顺学院学报(2021年4期)2021-09-16
奥秘(创新大赛)(2020年11期)2020-12-03
林业调查规划(2020年3期)2020-06-03
亚太传统医药(2019年3期)2019-04-18
中成药(2018年7期)2018-08-04
军民两用技术与产品(2018年11期)2018-07-16
现代农村科技(2018年6期)2018-01-18
