
虾青素保护线粒体和抑制氧化应激诱导的内皮祖细胞凋亡*
2014-05-15 00:52龚志刚丁世芳姜其钧付文波
医药导报 2014年6期
龚志刚,丁世芳,姜其钧,付文波
(1.南方医科大学研究生院,广州 510515;2.广州军区武汉总医院心内科,武汉 430070)
虾青素保护线粒体和抑制氧化应激诱导的内皮祖细胞凋亡*
龚志刚1,2,丁世芳2,姜其钧2,付文波2
(1.南方医科大学研究生院,广州 510515;2.广州军区武汉总医院心内科,武汉 430070)
目的 探讨虾青素对体外氧化应激诱导人外周血内皮祖细胞凋亡的影响及其机制。方法体外培养人外周血单核细胞源的内皮祖细胞,分为正常对照组、模型组(给予叔丁基过氧化氢100μmol·L-1)、预处理组(给予叔丁基过氧化氢100μmol·L-1+虾青素0.1,1.0,10.0 nmol·L-1预处理24 h)。噻唑蓝(MTT)法检测细胞存活率;4',6-二脒基-2-苯基吲哚(DAPI)染细胞检测细胞凋亡率;2',7'-二氯荧光黄双乙酸盐(DCFH-DA)法检测细胞内活性氧(ROS)水平;比色法检测半胱氨酸天冬氨酸蛋白酶(caspase-3)活性;阳离子脂质JC-1法测定线粒体膜电位。结果MTT检测发现,随着虾青素浓度的增加,内皮祖细胞的存活率也增加。与模型组[(48.5±4.3)%]比较,虾青素+叔丁基过氧化氢组细胞存活率明显升高[(57.6±8.2)%,(77.6±7.5)%,和(85.3±6.1)%,P<0.05];DAPI染细胞检测发现,预处理组凋亡细胞明显减少,模型组凋亡小体形成率(27.8±3.2)%,3个预处理组凋亡小体形成率分别为[(20.4±2.9)%,(14.9±1.7)%和(7.8±0.7)%,P<0.05];caspase-3活性检测表明,与模型组(0.345±0.018)比较,预处理组活性随虾青素剂量增大而依次降低[(0.291±0.013),(0.209±0.004),(0.169±0.013),P<0.05];DCHF-DA分子探针检测细胞内ROS发现,与模型组比较,预处理组细胞内ROS水平呈剂量依赖性降低,荧光强度明显减弱。JC-1法线粒体膜电位检测显示,与模型组比较,预处理组线粒体膜电位丢失被明显抑制。结论虾青素通过减少细胞内ROS,保护线粒体膜电位,下调caspase-3活性,最终起到抗氧化应激诱导的EPCs凋亡。
虾青素;氧化应激;内皮祖细胞;细胞凋亡;线粒体膜电位
内皮祖细胞(endothelial progenitor cells,EPCs)在外周循环血中的数量与功能是促进再内皮化,使其完整性得以维持的重要因素[1]。研究表明EPCs对氧化应激损伤敏感[2];活性氧(reactive oxygen species,ROS)水平升高是冠心病、缺血-再灌注损伤,以及经皮冠状动脉介入治疗损伤的共同特征[3],由此构成的引起内皮祖细胞损伤和凋亡的血管整体/局部氧化应激微环境,可能是患者外周血EPCs的数量及功能明显减少/受损的直接原因[4]。因此,探寻增强EPCs抗氧化应激能力,改善其数量和功能,可能是血管损伤后再内皮化治疗药物作用的新靶点[5]。虾青素(astaxanthin,ASX)对氧化应激引起的多种细胞如人神经干细胞[6],脐静脉内皮细胞[7],肾小球系膜细胞[8],视网膜细胞[9]等的凋亡具有显著保护作用。但ASX对EPCs是否具有抗氧化应激损伤引起凋亡的作用,目前笔者尚未见报道。笔者在本研究中利用体外培养人外周血单核细胞源的EPCs,叔丁基过氧化氢(tert-butyl hydroperoxide,tBHP)制作氧化应激损伤模型,观察并探讨ASX对tBHP诱导EPCs凋亡的保护作用及其机制。
1 材料与方法
1.1 药物与试剂 Histopaque1077淋巴细胞分离液(美国Amersham Biosciences公司);M199培养液、胎牛血清、细胞培养板(美国Gibico公司);胰酶(上海博光生物技术公司);硫氰酸荧光(fluorescein isothocyanate,FITC)标记荆豆凝集素I(FITC-UEA-I)、人纤维连接蛋白、ASX、叔丁基过氧化氢(tBHP)、四甲基偶氮唑盐(methylthiazolyldiphenyl-tetrazolium bromide,MTT)均购于美国Sigma公司;乙酰化低密度脂蛋白(DiI-Ac-LDL)购于美国Molecular Probe公司;重组人血管内皮生长因子(rh-VEGF165)、重组人碱性成纤维细胞生长因子(rh-b-FGF)均购于英国Pepro Tech公司;PE标记鼠抗人CD133单克隆抗体(德国Miltenyi Biotec公司);FITC标记鼠抗人CD34单克隆抗体(美国Southern Biotech公司);PE标记鼠抗人VEGF-R2单克隆抗体、Caspases活性检测试剂盒(美国R&D System公司);ROS检测试剂盒、线粒体膜电位检测试剂盒(JC-1)、DAPIApoptosis Detection试剂盒购自碧云天生物技术研究所。
1.2 人EPCs的分离、培养和鉴定 取健康志愿者外周血,以1∶2比例磷酸盐缓冲液(phosphate buffered solution,PBS)稀释后,缓慢移至Histopaque1077淋巴细胞分离液上方,400×g室温离心30 min,分离出中间单个核细胞层。将分离好的单核细胞用含有10% PBS、20%胎牛血清、rh-VEGF10 ng·mL-1、rh-b-FGF 10 ng·mL-1的M199培养液稀释成5×106·m L-1,放置于37℃,5%二氧化碳(CO2)孵箱中培养;培养到第4天,用PBS洗去非贴壁细胞,换培养液继续培养;取第7天的细胞,加入2.4 mg·L-1DiI-Ac-LDL和10 mg·L-1FITC-UEA-I于37℃孵育1 h后,用荧光显微镜检测双染色阳性细胞百分比。另取第7天的细胞,用0.25%胰蛋白酶消化,分别加入10 mg·L-1PE标记的VEGF-R2单克隆抗体、10 mg·L-1PE标记的CD133单克隆抗体、10 mg·L-1FITC标记的CD34抗体,4℃孵育30 min后,上流式细胞仪检测细胞表面标志物表达率。于第7天,加入无血清的培养液同步化细胞24 h,进行后续实验。
1.3 人EPCs的tBHP损伤模型的建立 同步化后的EPCs用0.25%胰酶消化,以每孔1×103的密度接种于96孔板,分别加入浓度为30,60,120,240μmol·L-1的tBHP干预6 h,加入MTT(5 g·L-1)10μL在37℃孵育4 h后吸去上清液,加入二甲亚砜100μL,振荡10 min溶解结晶,上酶标仪检测450 nm波长处的吸光度(A)值。根据细胞存活情况选择合适的tBHP作用浓度用于后续干预实验。细胞存活率(%)=(模型组A值/对照组A值)×100%。
1.4 ASX对正常和损伤细胞存活率的影响 待细胞长到铺满板底70%~80%时,预处理组加入终浓度分别为0.1,1.0,10.0 nmol·L-1的ASX溶液于37℃孵育24 h后,换为无血清MEM(minimum essential medium),再加终浓度为100μmol·L-1的tBHP溶液继续培养6 h后,MTT法检测细胞存活率。
1.5 细胞凋亡率的测定 药物处理好的细胞除去培养基,用PBS洗2次,binding buffer重悬并调整细胞数为1×106·mL-1,然后用4%多聚甲醛固定15 min,用DAPI(2.5μg·mL-1)染核20 min,在荧光显微镜下摄细胞图像,观察细胞核形态。荧光显微镜下,活细胞核呈弥散均匀荧光,细胞凋亡时,细胞核或细胞质内可见浓染致密的颗粒块状荧光,核出现不同程度的固缩,可见DNA荧光碎片(凋亡小体)。分别选取5个视野,并计数细胞100个,计算细胞凋亡小体形成率,细胞凋亡小体形成率(%)=凋亡小体细胞数/总的细胞数× 100%,取5次结果的平均值来评价细胞凋亡率。
1.6 测定caspase-3活性 按caspase-3 Colorimeric Assay Kit说明书进行。细胞1×106个溶于细胞溶解液50μL后,加入2×反应缓冲液50μL和酶底物(DEVD-pNA)5μL,37℃孵育1 h,用酶标仪在波长405 nm下测A值。
1.7 细胞内ROS水平测定 采用DCFH-DA检测细胞内ROS水平。96孔板,经药物处理后的各组EPCs,用10μmol·L-1DCFH-DA荧光探针的培养液于37℃细胞培养箱内孵育20 min,用无血清细胞培养液洗3次以去除探针,酶标仪检测DCF,激发波长488 nm,发射波长525 nm。DCF相对荧光强度(%)=处理组的荧光值/对照组的荧光值×100%。
1.8 细胞线粒体膜电位的测定 采用JC-1法。接种于96孔板,经药物处理后的各组EPCs,除去培养基, PBS洗2次,滴加JC-1工作液100μL,37℃,5%CO2孵箱中培养20 min,1×Incubation Buffer洗2次,于荧光显微镜下观察。JC-1是一种阳离子脂质荧光染料,作为检测线粒体跨膜电位指示剂。JC-1有单体和多聚体两种状态,低浓度时以单体形式存在,高浓度时以多聚体形式存在,两者的发射光谱不同,JC-1可透过正常细胞膜以单体状态聚集胞内,正常细胞线粒体的膜电位(ΔΨ)具有极性,JC-1依赖于ΔΨ的极性被迅速摄入线粒体内,并因浓度增高而在线粒体内形成多聚体,多聚体发射光为红色荧光;而在线粒体膜通透性改变,线粒体膜电位去极化时,JC-1从线粒体内释放,红光强度减弱,以单体的形式存在于胞质内发绿色荧光,根据这一特征检测线粒体膜电位的变化。
1.9 统计学方法 用SPSS13.0版统计分析软件进行分析,所有数据用均数±标准差(±s)表示,采用单因素方差分析及t检验,P<0.05为差异有统计学意义。
2 结果
2.1 EPCs的形态与鉴定 EPCs体外培养48 h后出现集落,4 d后多数集落已经形成,于第8天后加入DiI-Ac-LDL和FITC-UEA-I染色,其中90%细胞吞噬DiI-Ac-LDL(荧光显微镜下呈红色),及FITC-UEA-I (荧光显微镜下呈绿色)染色阳性。采用PE标记的鼠抗人CD133,FITC标记的鼠抗人CD34以及PE标记的鼠抗人VEGF-R2抗体于流式细胞仪检测细胞表面标志,结果显示CD133,CD34及VEGF-R2阳性率分别为(89.5±5.3)%,(66.3±6.3)%和(93.1±8.3)%。证实所分离培养的细胞为EPCs,见图1。

A.DiI-Ac-LDL染色;B.FITC-UEA-I染色;C.DiI-Ac-LDL+FITC-UEA-I染色图1 EPCs荧光染色鉴定图A.DiI-Ac-LDL staining;B.FITC-UEA-Istaining;C.DiI-Ac-LDL+FITC-UEA-IstainingFig.1 Fluorescent staining on EPCs
2.2 tBHP损伤EPCs模型的建立 MTT检测结果显示不同浓度tBHP作用EPCs 6 h后,细胞存活率表现为随药物浓度增加而降低的剂量依赖关系。当tBHP浓度在30μmol·L-1时,细胞存活率为(83.6±10.1)%, 60μmol·L-1时为(65.1±8.8)%,120μmol·L-1时为(34.7±13.2)%,至最大浓度240μmol·L-1时,细胞大部分死亡,细胞存活率为(6.3±4.6)%,与对照组(100%)比较,差异有统计学意义(P<0.05,n=6)。当tBHP浓度约为100μmol·L-1时,细胞存活率(48.5± 4.3)%(P<0.05,n=6),约维持在50%,后续实验选择该条件建立tBHP诱导EPCs的氧化损伤模型。见图2。
2.3 ASX对tBHP诱导的内皮祖细胞存活率下降的影响 MTT结果显示,预实验中给予浓度0.1,1.0, 10.0 nmol·L-1的ASX对EPCs的存活率没有影响。与模型组比较,各实验浓度的ASX呈剂量依赖性提高细胞存活率,当ASX的浓度达到10 nmol·L-1时,细胞存活率从(57.6±8.2)%升高至(85.3±6.1)%(P<0.05,n=6),显著提高EPCs存活率。
2.4 ASX对tBHP诱导EPCs凋亡的影响 应用DAPI染细胞检测ASX对tBHP引起EPCs凋亡的影响。tBHP 100μmol·L-1处理EPCs 6 h后,细胞出现典型的凋亡核形态学改变,与对照组比较,细胞染色质浓缩,核固缩明显,凋亡小体数目显著增加,ASX预处理24 h后,可显著改善细胞凋亡状况,随着作用浓度的增加,作用逐渐增强,ASX浓度在10.0 nmol·L-1时,改善最为显著。经过凋亡小体计算统计,正常对照组凋亡小体形成率为(4.8±0.7)%,100μmol·L-1tBHP损伤6 h组凋亡小体形成率(27.8±3.2)%,与正常对照组比较明显增多,差异有统计学意义(P<0.05,n=6), ASX在0.1,1.0,10.0 nmol·L-1预处理24 h后再加100μmol·L-1tBHP处理6 h,凋亡小体形成率分别为(20.4±2.9)%,(14.9±1.7)%和(7.8±0.7)%(P<0.05,n=6),可见ASX对tBHP诱导的细胞凋亡具有明显的保护作用,见图3。
2.5 ASX对tBHP诱导EPCs caspase-3活性的影响应用紫外分光光度法检测EPCs caspase-3活性,结果显示:与正常对照组(0.096±0.014)比较,模型组caspase-3活性(0.345±0.018)显著升高(P<0.05,n= 6);而经ASX预处理24 h后,与模型组比较,随着ASX剂量从0.1,1.0,10.0 nmol·L-1增加而caspase-3活性依次下降[(0.291±0.013),(0.291±0.013),(0.169± 0.013),P<0.05,n=6],提示ASX可通过调节caspase-3活性抑制tBHP诱导的EPCs细胞凋亡。

A.对照组;B.模型组;C.预处理组图2 3组内皮祖细胞存活检测A.control group;B.model group;C.pretreatment groupFig.2 Detection on cell viability of three groups of EPCs

A.对照组;B.模型组;C.预处理组图3 3组内皮祖细胞凋亡的检测A.control group;B.model group;C.pretreatment groupFig.3 Detection on cell apoptosis of three groups of EPCs
2.6 ASX对细胞内ROS的影响 tBHP渗透进入细胞,迅速转化为叔丁氧基,并进而促进细胞内ROS的形成。DCHF-DA是检测细胞内ROS的分子探针,其本身无荧光,当它进入细胞与细胞内ROS结合时,产生一种荧光物质DCF,DCF的荧光强度反映细胞内ROS水平。本实验检测结果显示,100μmol·L-1tBHP损伤组,EPC细胞内ROS水平显著增加,荧光强度增大;而经ASX(0.1,1.0,10.0 nmol·L-1)预孵育组, EPCs细胞内ROS水平呈剂量依赖性降低,ASX浓度在10 nmol·L-1时,ROS水平降低最为显著,荧光强度较损伤模型组明显减弱,见图4。
2.7 EPCs线粒体膜电位检测 本实验荧光显微镜下观察结果显示正常对照组为黄绿色,模型组线粒体膜电位丢失,绿色荧光强度变强,而经10 nmol·L-1ASX预处理24 h后,其线粒体膜电位丢失得以明显抑制,呈现黄绿素荧光,但较对照组黄绿色稍偏绿;说明ASX能抑制tBHP引起的EPCs线粒体膜电位去极化,从而提高线粒体膜电位水平,进而对线粒体膜电位丢失促发的EPCs凋亡具有一定的保护作用,见图5。
3 讨论
ROS是细胞凋亡中的第二信使[10]。研究证实,氧化还原失衡引发的ROS过度升高(氧化应激)可加重内皮祖细胞凋亡和损伤,造成其数量和功能下降,严重影响血管内皮修复,引起再内皮化不全或延迟,导致临床心血管不良事件的发生。
ASX是一种酮式类胡萝卜素,源自雨生红球藻类受到较长时间环境胁迫时,细胞内产生的一种抵御不良环境的长期分子机制,具有多种生物活性,其中最重要的是其极高的抗氧化活性,可应对细胞内氧化应激造成的损伤,使细胞渡过难关[11],tBHP是用于构建细胞氧化损伤的常用试剂,其化学性质与过氧化氢(H2O2)相似,但更稳定,不易降解。tBHP能模拟氧化应激损伤,直接破坏膜结构,最终导致细胞或线粒体膜的破坏[12]。

A.对照组;B.模型组;C.预处理组图4 3组内皮祖细胞内ROS的检测A.control group;B.model group;C.pretreatment groupFig.4 Detection on ROS in three groups of EPCs
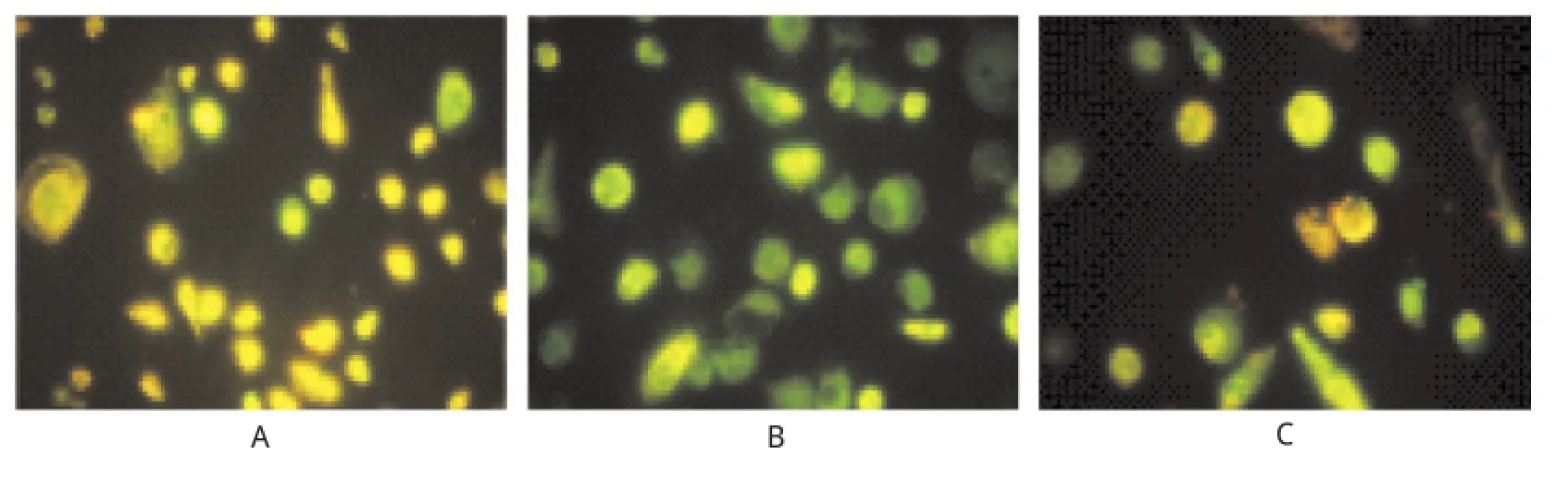
A.对照组;B.模型组;C.预处理组图5 3组内皮祖细胞线粒体膜电位的检测A.control group;B.model group;C.pretreatment groupFig.5 Detection on mitochondrialmembrane potential in three groups of EPCs
细胞凋亡是多基因、多种分子参与的复杂生命过程。研究发现,caspase-3是细胞凋亡发生和调控机制中的重要分子,是凋亡途径下游进行底物酶解的关键蛋白酶,caspase-3的激活是凋亡发生的最早期事件,它对染色质固缩、DNA片段化起重要的执行作用[13]。本研究通过DAPI检测发现tBHP可明显的引起内皮祖细胞的凋亡,单纯ASX对内皮祖细胞无损伤作用,ASX对tBHP诱导的内皮祖细胞凋亡具有明显的保护作用。在ASX抑制EPCs的凋亡中发现ASX对caspase-3的活性有下调效应,在10.0 nmol·L-1水平能显著抑制其激活。
研究发现线粒体在细胞凋亡过程中起着枢纽作用,线粒体膜电位的下降被认为是细胞凋亡级联反应中的早期表现,线粒体膜电位一旦破坏,可导致线粒体外膜的破裂,各种凋亡因子会随之释放,导致细胞凋亡的发生[14]。本实验发现,tBHP损伤组细胞线粒体膜电位明显低于空白对照组,而ASX保护组的细胞线粒体膜电位较损伤组有明显的提高,可见tBHP诱导的氧化应激损伤降低了EPCs线粒体膜电位,启动了线粒体途径细胞凋亡,而ASX能明显提高EPCs线粒体膜电位,抑制tBHP诱导的线粒体途径细胞凋亡。
综上所述,本研究显示ASX通过减少细胞内ROS,保护线粒体膜电位,下调caspase-3活性,最终起到抗氧化应激诱导的EPCs凋亡。本研究为ASX抗氧化应激损伤,改善EPCs数量和功能,促进损伤血管再内皮化提供了实验依据。
[1] BALAJI S,KING A,KESWANI S.The role of endothelial progenitor cells in postnatal vasculogenesis[J].AdvWound Care,2013,2(6):283-295.
[2] INGRAM D A,KRIER T R,MEAD L E,et al.Clonogenic endothelial progenitor cells are sensitive to oxidative stress [J].Stem Cells,2007,25(2):297-304.
[3] TURGEON J,DUSSAULT S,HADDAD P,et al.Probucol and antioxidant vitamins rescue ischemia-induced neovascularization in mice exposed to cigarette smoke: potential role of endothelial progenitor cells[J]. Atherosclerosis,2010,208(2):342-349.
[4] WERNER N,KOSIOL S,SCHIEGL T,et al.Circulating endothelial progenitor cells and cardiovascular outcomes [J].N Engl JMed,2005,353(10):999-1007.
[5] IMANISHIT,TSUJIOKA H,AKASAKA T.Endothelial progenitor cells dysfunction and senescence:contribution to oxidative stress[J].Curr Cardiol Rev,2008,4(4):275-286.
[6] KIM J H,CHOIW,LEE J H,et al.Astaxanthin inhibits H2O2-mediated apoptotic cell death in mouse neural progenitor cells viamodulation of P38 and MEK signaling pathways[J].Microb Biotech,2009,19(11):1355-1363.
[7] ADLURIR S,THIRUNAVUKKARASU M,ZHAN L,et al. Cardioprotective efficacy of a novel antioxidantmix Vitae-Pro against ex vivo myocardial ischemia-reperfusion injury [J].Cell Biochem Biophys,2013,67(2):281-286.
[8] MANABE E,HANDA O,NAITO Y,et al.Astaxanthin protectsmesangial cells from hyperglycemia-induced oxidative signaling[J].J Cell Biochem,2008,103(6):1925-1937.
[9] DONG L Y,JIN J,LU G,et al.Astaxanthin attenuates the apoptosis of retinal ganglion cells in db/db mice by inhibition of oxidative stress[J].Mar Drugs,2013,11(3): 960-974.
[10] DERNBACH E,URBICH C,BRANDES R P,et al.Antioxidative stress-associated genes in circulating progenitor cells:evidence for enhanced resistance against oxidative stress[J].Blood,2004,104(12):3591-3597.
[11] PASHKOW F J,WATUMULLDG,CAMPBELLC L.Astaxanthin:a novel potential treatment for oxidative stress and inflammation in cardiovascular disease[J].Am J Cardiol, 2008,101(10A):58-68.
[12] HAIDARAK,MOREL I,ABALEA V,et al.Mechanism of tertbutylhydroperoxide induced apoptosis in rat hepatocytes:involvement of mitochondria and endoplasmic reticulum[J].Biochim Biophys Acta,2002,1542(1):173-185.
[13] CASE A,INGRAM D A,HANELINE L S.Oxidative stress impairs endothelial progenitor cell function[J]. Antioxidants Redox Signaling,2008,10(11):1895-1907.
[14] FU D,CHEN T,SZETO H,et al.Mitochondrial targeted antioxidant peptide ameliorateshypertensive cardiomyopathy [J].J Am Coll Cardiol,2011,58(1):73-82.
DOI 10.3870/yydb.2014.06.005
Astaxanthin Inhibits Endothelial Progenitor Cell Apoptosis Induced by Oxidative Stress via Mitochondria-targeted Protective Mechanism
GONG Zhi-gang1,2,DING Shi-fang2,JIANG Qi-jun2,FU Wen-bo2
(1.Graduate School of Southern Medical University,Guangzhou 510515,China;2.Department of Cardiology,Wuhan General Hospital of Guangzhou Command,Wuhan 430070,China)
ObjectiveTo investigate the effect of astaxanthin(ASX)on endothelial progenitor cells(EPCs)injury induced by oxidative stressin vitroand to explore its underlyingmechanism.MethodsCultured EPCs isolated from peripheral blood were randomly divided into5 groups:normal control,model group[tert-butyl hydroperoxide(tBHP)100μmol·L-1],and ASX+tBHP groups(the cellswere preconditioned with ASX 0.1,1.0,and 10.0 nmol·L-1,respectively).The cell viability was measured by MT Tmethod.The level of reactive oxygen species(ROS)was determined by DCFH-DA method.The changes of mitochondrialmembrane potential(MMP)and apoptosis ratio were detected by JC-1 method and DA PImethod,respectively. caspase-3 activity changes of EPCs were detected.ResultsThe cell viability of EPCs was improved with the increasing concentration of ASX.Compared with themodel group[(48.5±4.3)%],0.1,1.0,10.0 nmol·L-1ASX significantly increased the cell viabilities[(57.6±8.2)%,(77.6±7.5)%,and(85.3±6.1)%,P<0.05].The results of DAPIstaining revealed that ASX pretreatment could significantly reduce the apoptotic rate of EPCs.The apoptotic rate of the model group was(27.8± 3.2)%,while that of ASX+tBHP groupswas[(20.4±2.9)%,(14.9±1.7)%,and(7.8±0.7)%,P<0.05],respectively.The data from caspase-3 activity assay indicated that ASX precondition could also remarkably decrease the caspase-3 activity forEPCs.The caspase-3 activity of the model group was(0.345±0.018),while that of the ASX+tBHP group were[(0.291± 0.013),(0.209±0.004),and(0.169±0.013),P<0.05],respectively.In addition,treatmentwith tBHP resulted in an increase of DCF fluorescence,while ASX precondition could decrease the DCF fluorescence,which suggested the accumulation of intercellular ROS for EPCs.Injury of michondrialmembrane resulted in the loss of mitochondrial membrane potential(MMP). The MMP detected by JC-1 method revealed that compared with model group,pretreatment of ASX inversed the reduction of MMP.ConclusionAstaxanthin inhibits endothelial progenitor cell apoptosis induced by oxidative stress through inhibiting ROS production,improving themitochondrial function and down-regulating caspase-3 activity.
Astaxanthin;Oxidative stress;Endothelial progenitor cell;Cell apoptosis;Mitochondrialmembrane potential
R966
A
1004-0781(2014)06-0712-06
2013-10-03
2013-12-18
*湖北省自然科学基金项目(2012FFB06803)
龚志刚(1971-),男,湖北随州人,副主任医师,博士,从事心内科临床工作。电话:(0)13397199595,E-mail:13397199595@189.cn。
丁世芳(1957-),女,山东招远人,主任医师,教授,从事心内科临床研究工作。电话:027-68879415,E-mail:dingshifangmd@yahoo.com.cn。
猜你喜欢
世界科学技术-中医药现代化(2021年7期)2021-11-04
教育教学论坛(2019年19期)2019-06-17
安徽医科大学学报(2015年9期)2015-12-16
医学研究杂志(2015年11期)2015-06-10
中国药理学通报(2014年2期)2014-05-09
中国医学科学院学报(2013年3期)2013-03-11
中国民间疗法(2013年8期)2013-01-25
中国医学科学院学报(2012年3期)2012-03-25
