
江西武夷山南方铁杉群落分类及更新能力评价
2014-12-14 02:50杨清培钟安建金志农程松林裘利洪施建敏郭英荣陈伏生邹思成
江西农业大学学报 2014年6期
杨清培,钟安建,金志农,程松林,裘利洪,施建敏,郭英荣,陈伏生,邹思成
(1.江西农业大学 江西省竹子种质资源与利用重点实验室,江西 南昌 330045;2.江西省九江市庐山区林业局,江西 九江 332000;3.江西农业大学 江西省鄱阳湖流域农业生态工程技术研究中心,江西 南昌 330045;4.江西武夷山国家级自然保护区管理局,江西 南昌 330045)
南方铁杉(Tsuga tchekiangensis)又名浙江铁杉,系松科铁杉属常绿高大乔木,是中国特有第三纪残遗植物之一,在松科植物起源与迁移方面有很高科研价值[1]。同时,南方铁杉材质优良、纹理细致、坚实耐用,可广泛应用于建筑、航空、造船和家具[2],但长期以来南方铁杉原生植被遭受不断破坏,造成生存竞争力降低、种群数量不断减少,适生面积缩小、空间分布斑块化,被列为国家三级重点保护植物[3-4]。
近年来,在南方铁杉生长规律[5]、种群动态[6]、群落结构[7]、菌根生态[8]与育苗技术[9-10]等方面已开展了大量的研究工作,这对了解它的生物学特性及保护对策提供了重要参考。然而有关南方铁杉更新能力的问题,却存在较大争议。冯祥麟等[7]认为贵阳高坡、广西猫儿山南方铁杉自然繁殖能力差,难以维持种群的更新,郭连金[11]与黄宪刚[6]还认为广西猫儿山和江西武夷山南方铁杉种群早在100 年前就开始衰退,杜道林[12]还预言600 年后南方铁杉会从贵州茂兰喀斯特山地完全消失,但张志祥[3]却认为南方铁杉种群结构虽存在一定的波动,但仍能进行自然更新。那么南方铁杉的更新能力到底怎样,关键制约因素是什么呢?
任何物种都不是孤立存在,它必然与其物种相互作用组成群落[13],因此任何种群现状都是它与其伴生物种长期协同进化的结果[14]。要了解南方铁杉种群的更新状况,必须了解它存在的群落类型及环境状况。然而,尽管南方铁杉分布范围广泛,分布格局却呈现不连续分布,多散生于落叶阔叶林或常绿落叶阔叶混交林中[2],因此,有关南方铁杉群落分类的研究较少,这必然会影响南方铁杉更新能力评价及保护政策的实施。
种群更新能力是指植物种群通过参与群落竞争,获得新个体补充的能力及种群延续能力,自然更新失败是植物种群衰退、消失的直接原因[15]。因此研究植物自然更新是濒危植物保护生物学的主要研究内容之一[16-17]。然而遗憾的是,目前对植物种群更新能力评价主要是通过种群年龄结构[18]、种子库[19]、幼苗数量与质量[20]等指标来反映,这些指标单独使用都难以满足种群更新能力的综合评价。为此,很有必要构建一个量化指标来评价南方铁杉在不同群落中的更新能力。
江西武夷山自然保护区内保存有南方铁杉天然林1 560 hm2,是全国为数不多的较大局域种群[21]。本研究在全面群落调查的基本上,先运用二元指示种分析(TWINSPAN)和去趋势典范对应分析(DCCA)等方法[22-23],对江西武夷山自然保护区南方铁杉群落进行了数量划分与环境解释,然后对南方铁杉更新能力进行了定量评价。试图回答以下3 个问题:(1)南方铁杉群落有哪些基本类型?(2)南方铁杉不同群落中的更新能力怎样?(3)影响南方铁杉群落分布与种群更新的环境因子有哪些?通过回答以上3 个问题,以期为南方铁杉群落多样性维持机制、南方铁杉种群就地(迁地)保护提供依据。
1 研究区自然概况
江西武夷山国家级自然保护区位于江西省上饶市铅山县,地处武夷山脉北段的西北坡,地理坐标范围为东经117°39'30″~117°55'47″,北纬27°48'11″~28°00'35″。保护区总面积16 007 hm2,其中核心区4 835 hm2,缓冲区2 021 hm2,实验区9 151 hm2,其主峰黄岗山,海拔2 158 m,是我国华东大陆最高峰,该区植被垂直分布明显。在保护区内保存有1 560 hm2完整的南方铁杉天然林资源,为该地区中山温性针叶林的典型代表[21]。在前期全面踏查的基础上,在海拔1 400~1 900 m 地段选择不同坡向、不同坡位、不同类型南方铁杉群落进行调查。
2 研究方法
2.1 样地设置与植被调查
野外调查于2010 年11 月和2011 年7—8 月份进行。在前期调查的基本上,按海拔、坡位、群落类型、更新状况等,设置30 个固定样地,本文将有南方铁杉出现的群落统称为南方铁杉群落,而不管其在群落(或群落层次中)中的优势度。样地面积20 m×30 m,并将其划分成6 个10 m×10 m 小样方,同时每小样方内嵌套设置1 个5 m×5 m 的灌木样方和1 个1 m×1m 的草本样方。对乔木、灌木进行每木检尺,记录种类、胸径、树高等。对草本记录种类、高度、盖度、多度。同时,用手持GPS 测量海拔、经纬度;用罗盘仪测量坡度、坡向,另外记录样地的坡位、主林层(灌木层)郁闭度、凋落物厚度、土壤厚度等环境因子。
2.2 土壤与分析
2011 年8 月,每10 m×10 m 小样方内采取3 个土壤样品(0~20 cm 表层土壤),混合装入布袋,组成待测土样,带回实验室风干、研磨、过筛,进行化学分析。铵态氮(NH4+-N)测定采用靛酚蓝比色法;硝态氮(NO3--N)测定采用镀铜镉还原-重氮化偶合比色法;全氮(STN)采用半微量凯氏法;有机碳(SOC)采用重铬酸钾外加热氧化法;土壤容重采用环刀法;土壤含水率采用烘干法(105 ℃,12 h);全磷用NaOH 熔融-钼锑抗比色法,速效P 用NaHCO3提取-钼锑抗比色法[24]。
2.3 群落分类与排序
根据样方调查数据,按文献[25]分别计算各样地乔木层、灌木层和草本层中所有物种的重要值(IV),剔除偶见种,选定重要值不小于5%的120 个种,形成30×120 的物种矩阵,同时以15 个环境因子构成30×15 总环境因子矩阵,另外还构建30×5 空间因子矩阵、30×10 土壤因子矩阵两个次矩阵用于方差分解分析。最后采用了生态学软件PCORD 5.0 的TWINSPAN 和CANOCO 5.0 的DCCA 群落分类或排序[26-27]。
2.4 种群更新能力
任何植物种群无时不在与群落中其他种群发生竞争[13],因此更新能力应该包括种群大小、年龄结构和相对多度(密度)。种群大小反映种群发展能力、年龄结构预示种群发展趋势,相对多度(密度)反映种群在群落中竞争的能力。为比较同一种群在不同群落中更新能力,本文假定以上3 者贡献相同,并试采用下列式中进行计算:

式(1)中,RP 代表种群更新能力,取值[0~1];UAS 种群年龄结构归一化值,种群结构赋值为:增长型种群4;稳定种群3;波动种群2;衰退种群1;没有出现0;UAB 代表种群多度(或密度)归一化值;URA代表相对多度(某种群个体数与群落全部个体数的比)归一化值。归一化处理采用公式:Xi’=(Xi-Xmin)/(Xmax-Xmin)进行,其中Xi分别表示该种群在群落i 中的多度(密度),相对多度和年龄结构赋值。Xmin、Xmax分别表示该种群全部群落中的多度(密度),相对多度和年龄结构赋值的最小值与最大值。
3 结果与分析
3.1 南方铁杉群落物种组成与垂直结构
30 个样地(18 000 m2),共调查到维管植物210 种,隶属于68 科112 属。其中被子植物60 科;裸子植物3 科;蕨类植物5 科。其中蔷薇科最多,有6 属15 个种,其次是山茶科6 属14 种,山矾科1 属11种,杜鹃花科3 属10 种,冬青科1 属10 种,樟科3 属9 种,禾本科8 属9 种。另外,还孕育有南方红豆杉(Taxus mairei)、天目紫茎(Stewartia gemmata)、银钟花(Halesia macgregorii)等珍稀植物和柳杉(Cryptomeria fortunei)、大血藤(Sargentodoxa cuneata)等古老孑遗植物[28],表明武夷山南方铁杉群落的物种比较丰富,而且起源也较古老。同时,南方铁杉群落层次分化明显,乔木、灌木和草本发育完整,甚至部分群落还有丰富的层间植物。
3.2 南方铁杉群落TWINSPAN 数量分类
采用TWINSPAN 程序将30 个武夷山南方铁杉样地划分为11 个群落类型(图1),它们分别是:Ⅰ.多脉青冈-箬竹-荩草群落(Ass.Cyclobalanopsis multinervis-Indocalamus tessellatus-Arthraxon hispidus)。乔木主要种类有多脉青冈、皖闽八角等常绿树种;灌木层箬竹入侵严重,未被侵占地方有皖闽八角(Illiciumminwanense)等常绿植物,天目紫茎、中华石楠(Photinia beauverdiana)等落叶植物,草本层种类较少。

图1 30 个南方铁杉样地的TWINSPAN 分类树状图。D 表示分级水平;N 表示样地数量Fig.1 Dendrogram of the TWINSPAN classification of 30 plots of Tsuga tchekiangensis.D indicates the level of division;N indicates the number of plots
Ⅱ.光皮桦+南方铁杉-江南山柳+南方铁杉-苔草群落(Ass.Betula luminifera+Tsuga tchekiangensis-Clethra cavaleriei+Tsuga tchekiangensis-Carex chinensis)。乔木层主要是光皮桦(Betula luminifera)、尾叶樱(Cerasus dielsiana)等落叶树种伴生,灌木层主要有江南山柳(Clethra cavaleriei)、吊钟花(Enkianthus quinqueflorus)等落叶植物。
Ⅲ.雷公鹅耳枥+南方铁杉-南方铁杉-中华苔草群落(Ass.Cyclobalanopsis multinervis+Tsuga tchekiangensis-Tsuga tchekiangensis-Carex chinensis)。乔木层有雷公鹅耳枥、缺萼枫香(Liquidambar acalycina)等落叶树种,也有南方铁杉、多脉青冈等常绿乔木;灌木层主要是鹿角杜鹃、微毛山矾(Symplocos wikstroemiifolia)等常绿植物,落叶植物仅有天目紫茎等少数种类,总盖度为30%~50%。
Ⅳ.多脉青冈+南方铁杉-鹿角杜鹃-瘤足蕨群落(Ass.Cyclobalanopsis multinervis+Tsuga tchekiangensis-Rhododendron latoucheae-Plagiogyria adnata)。乔木层主要有南方铁杉、多脉青冈等常绿乔木,也有天目紫茎、缺萼枫香等落叶乔木;灌木层主要有鹿角杜鹃、格药柃(Eurya muricata)等常绿灌木,并开始受毛秆箭竹入侵。
Ⅴ.南方铁杉+多脉青冈-毛秆箭竹-苔草群落(Ass.Tsuga tchekiangensis+Cyclobalanopsis multinervis-Sinarundinaria hirticaulis-Carex chinensis)。乔木层主要是南方铁杉、多脉青冈、皖闽八角等常绿乔木;灌木层还有鹿角杜鹃、格药柃等常绿植物,毛秆箭竹入侵严重,总盖度为50%~60%。
Ⅵ.多脉青冈+南方铁杉-毛秆箭竹-灯芯草群丛(Ass.Tsuga tchekiangensis+Cyclobalanopsis multinervis-Sinarundinaria hirticaulis-Juncus effusus)。乔木层主要有多脉青冈、南方铁杉等常绿种类,也有光皮桦、秀丽槭(Acer elegantulum)等落叶种类;灌木层毛秆箭竹入侵严重,盖度为80%。
Ⅶ.黄山松+南方铁杉-毛秆箭竹-苔草群丛(Ass.Pinus taiwanensis+Tsuga tchekiangensis-Sinarundinaria hirticaulis-Carex chinensis)。乔木层主要有黄山松、南方铁杉、多脉青冈等常绿种类;灌木层主要有鹿角杜鹃、格药柃等常绿灌木,毛秆箭竹入侵严重,总盖度为50%~60%。
Ⅷ.南方铁杉+黄山松-毛秆箭竹-黄背草群丛(Ass.Tsuga tchekiangensis+Pinus taiwanensis-Sinarundinaria hirticaulis-Themeda japonica)。乔木层主要有南方铁杉、黄山松、柳杉等常绿种类;灌木层为马银花(Rhododendron ovatum)、岩柃(Eurya saxicola)常绿灌木较多,并有毛秆箭竹入侵,盖度为20%~40%。
Ⅸ.黄山松+南方铁杉-满山红+吊钟花-苔草群丛(Ass.Pinus taiwanensis+Tsuga tchekiangensis-Rhododendron mariesii+Enkianthus quinqueflorus-Carex chinensis)。乔木层有黄山松(Pinus taiwanensis)、南方铁杉等常绿乔木;灌木层有满山红、吊钟花等落叶种类,也有鹿角杜鹃、岩柃等常绿种类,总盖度为40%~50%。
Ⅹ.南方铁杉-满山红-苔草群丛(Ass.Tsuga tchekiangensis-Rhododendron mariesii-Carex chinensis)。乔木层有南方铁杉、黄山松等常绿乔木,灌木层主要有满山红、山柳(Clethra barbinervis)等落叶树种,常绿种类只有岩柃等少数种类。
Ⅺ.豆梨-箬竹+南方铁杉-苔草群丛(Ass.Pyrus calleryana-Indocalamus tessellatus+Tsuga tchekiangensis-Carex chinensis)。乔木层主要种类有豆梨、多脉青冈等,灌木层主要有箬竹、南方铁杉、岩柃、白檀(Symplocos paniculata)、饭汤子(Viburnum setigerum)等;草本层盖度为5%~10%,种类较少。
3.3 DCCA 排序及对分类结果的解释
3.3.1 DCCA 排序 南方铁杉群落30 个样地DCCA 排序结果(图2)。从图2 可知,海拔、土壤厚度、无机氮与Axis 1 显著正相关;有效磷、总磷与Axis 2 呈显著正相关。所以从左上角到右下角,表示海拔、坡位逐渐升高,土壤磷素逐渐减小,而从左下到右上,土壤厚度逐渐加深,土壤氮含量越丰富。
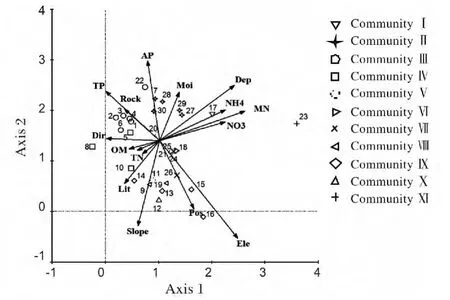
图2 武夷山南方铁杉群落的DCCA 排序图(1-30 代表样地号)Fig.2 Two dimensional scatter plots of DCCA ordination for 30 plots(The number from 1-30 in the map represents the code of the plots)
由图2 还可看出群落空间分布及生态关系,群落类型Ⅰ(样地17)主要分布在土层较厚、无机氮丰富的中低海拔地段;群落类型Ⅱ(样地7,27,28,29,30)主要分布在海拔较低,湿度较大,土壤有效磷较丰富的地段;群落类型Ⅲ(1,2,3,4,6,22)主要分布在海拔较低、岩石裸露率较高、总磷含量高的地段;群落类型Ⅳ(样地5,8,10)主要分布中低海拔的阳坡;群落类型Ⅴ(样地11,20,21,25)、群落类型Ⅵ(样地18)、群落类型Ⅶ(样地24,26)主要位于各项条件中等地段;群落类型Ⅷ(样地9,19)主要分布在中海拔、坡度较大的地段;群落类型Ⅸ(13,15,16),Ⅹ(样地12)主要分布在海拔较高的上坡或山脊,而群落类型Ⅺ(样地23)离其他群落最远,喜欢在无机氮较为丰富的地方。可见DCCA 排序结果与TWINSPAN分类结果具有较好的吻合度。
当然,从群落间生态关系来看,除类型Ⅺ与其它关系稍远外,其它类型间关系较近,一方面说明本区南方铁杉群落间生态关系较近,不论在时间或空间是南方铁杉都呈连续分布;另一方面也说明南方铁杉在该区适应性较强,完全适合于各海拔、坡度、坡向和各种土壤条件。
3.3.2 环境因子方差分解 南方铁杉环境因子方差分解分析,土壤因子和空间因子对群落分布都具有一定影响,但土壤因子比空间要素的影响力大,前者单独影响力为37.73 %,后者为15.48 %,二者偶合效应为7.65 %(图3),表明空间因子对土壤环境的能流和物流有着重新分配和控制能力[29]。
3.4 种群更新能力及环境解释
3.4.1 不同群落南方铁杉更新能力评价 南方铁杉更新能力在不同类型群落中评价(表3)。由表3 可知,南方铁杉在群落类型Ⅱ中更新能力最强,样地27、7、30、28 分别排名第1,3,6,7;在类型Ⅸ中更新次之,样地14、16,15、13分别排名第2,4,9,11;而在群落类型Ⅵ中,更新较为困难,样地20、11、21,更新能力排名第30、27、23。

图3 南方铁杉群落格局的土壤环境、空间因子方差分解及解释能力Fig.3 Variation partioning of Tsuga tchekiangensisa communities and environmental interpretation
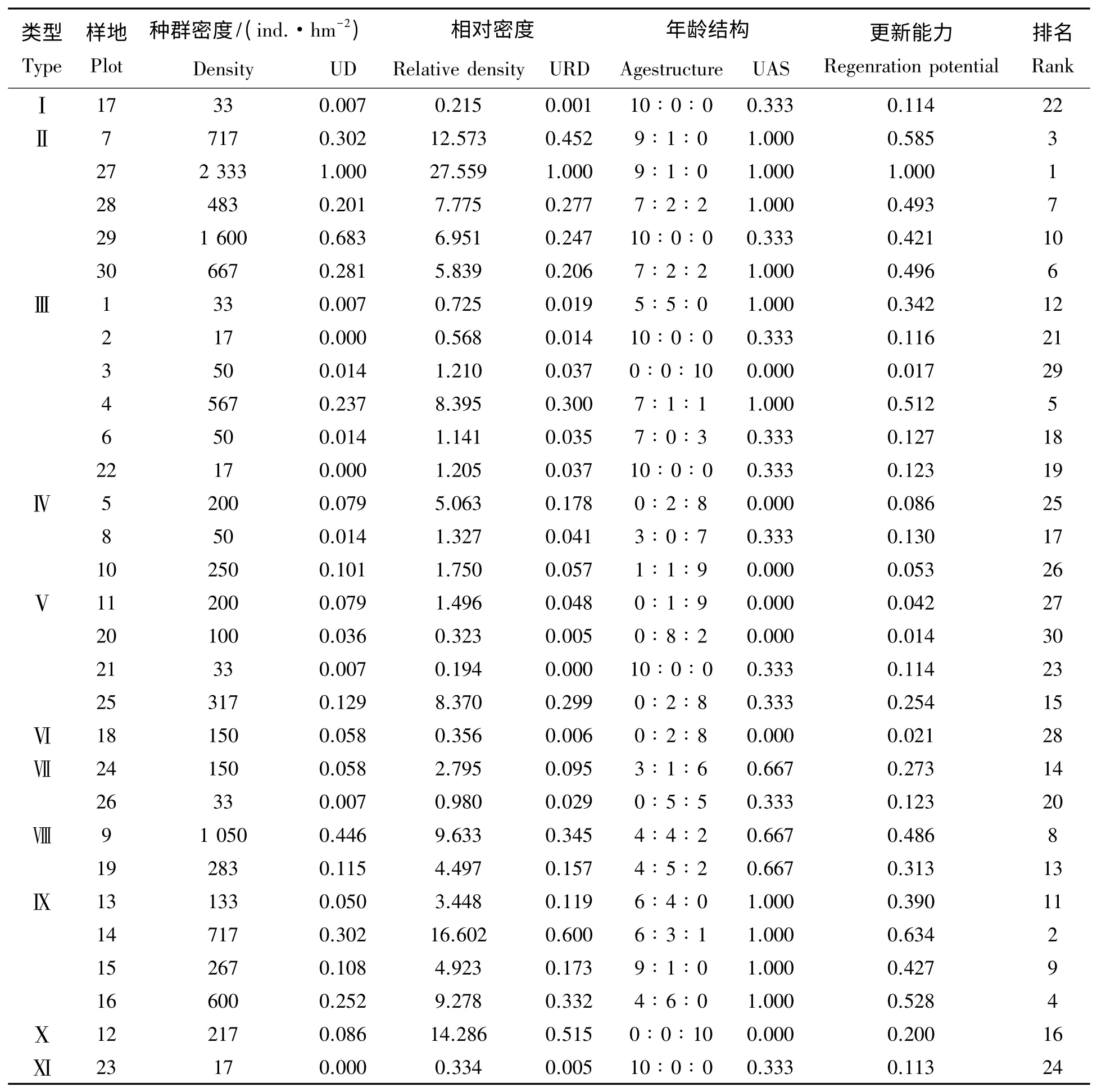
表1 南方铁杉种群在不同群落中更新能力Tab.1 Natural regeneration capacity of Tsuga tchekiangensis in different types of community
3.4.2 南方铁杉自然更新的影响因子 南方铁杉更新能力与群落空间位置、土壤养分状况(除总磷与有效磷外)关系不明显,而与群落组成与结构特征关系密切(表4)。表4 可知,南方铁杉更新能力与群落生物多样性、乔木层落叶树重要值、灌木层落叶树重要值呈明显正相关,而与群落优势度、乔木层常绿树种重要值、灌木层常绿树种重要值呈显著负相关,尤其是灌木层组成与盖度对南方铁杉更新影响最大。如在群落类型Ⅱ(样地27、7、30、28)中,乔木层植物主要是枫香、雷公鹅耳枥、光皮桦等落叶阔叶树,灌木层中也主要是满山红、天目紫茎、江南山柳等落叶植物,林分透光性好,其更新能力排名分别为第1,3,6,7。即使乔木层是南方铁杉、黄山松等常绿树种,只要林下主要是落叶植物,则南方铁杉更新也较好;如在群落类型Ⅸ的样地14、16,15,乔木层是黄山松、南方铁杉,但灌木层主要是江南山柳、满山红、吊钟花等,更新能力排名分别为第2,4,9。
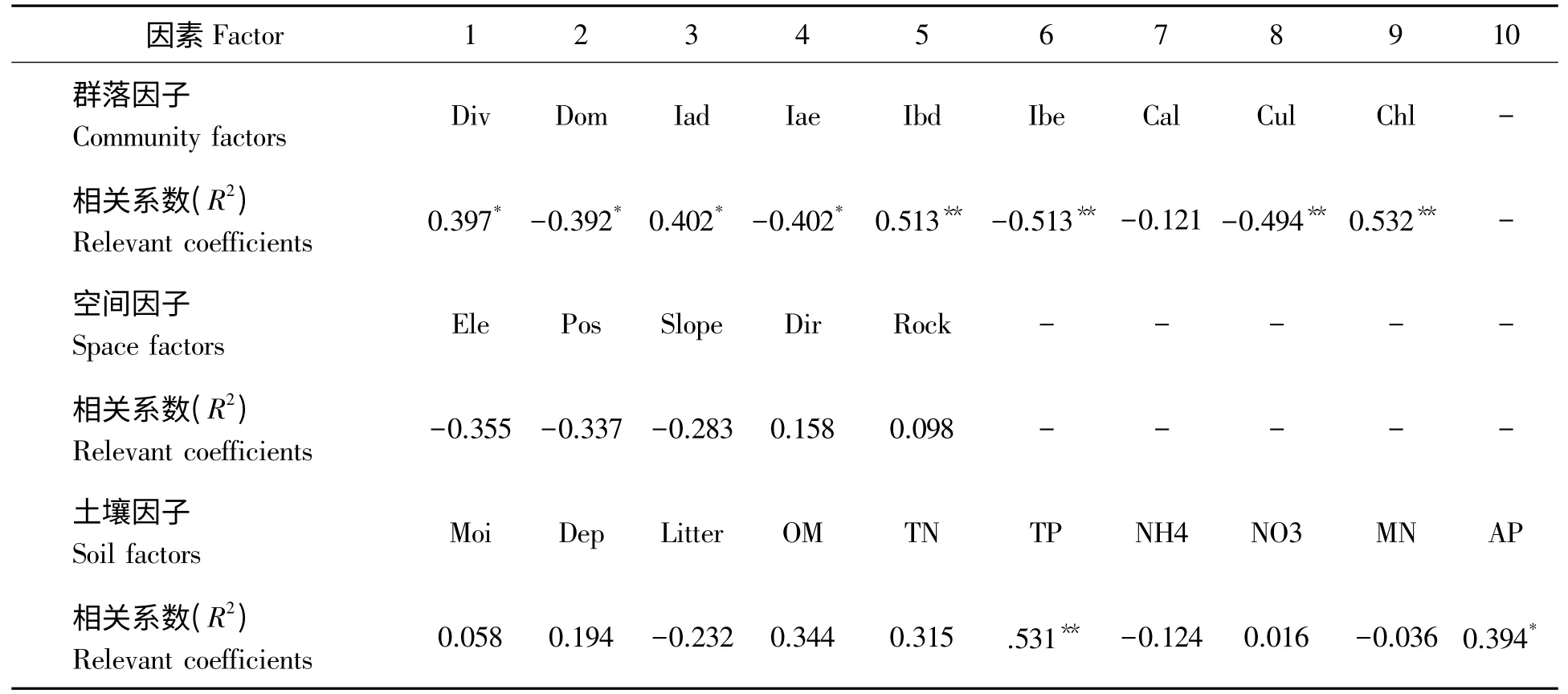
表2 南方铁杉自然更新能力与群落特征及环境因子的关系Tab.2 The relationship of Tsuga tchekiangensis natural regeneration capacity and communities,environmental factors
相反,如灌木层常绿树种占优势、林下盖度增加时,南方铁杉更新变得困难,如在群落类型Ⅲ的样地22、2 中,虽乔木层是雷公鹅耳枥、光皮桦,但林下优势种却为多脉青冈、多穗石栎、黄丹木姜子(Litsea elongata)等,其更新能力排名第19、21;当然,如乔木层、灌木层全是常绿树种占优势时,南方铁杉更新则非常困难,如群落类型Ⅵ的样地21、11、20,乔木层中主要是多脉青冈、皖闽八角等常绿树种,灌木层中也主要是鹿角杜鹃、皖闽八角、薄叶山矾、岩柃等常绿树种,更新能力排名第23、27、30,尤其是毛秆箭竹大量入侵,毛秆箭竹占据绝对优势,不仅南方铁杉更新困难,而且整个群落物种多样性大大降低。说明南方铁杉在由落叶阔叶树(光皮桦、枫香、雷公鹅耳枥、山樱花Cerasus serrulata)组成的群落中更新良好,但随着多脉青冈、皖闽八角、多穗石栎、鹿角杜鹃等常绿阔叶植物的进入,南方铁杉更新变得困难,尤其是林下受到毛秆箭竹、箬竹等灌木性竹类植物大量入侵时,灌木层郁闭度增加、地下竞争激烈,其更新能力大大下降,甚至完全不能更新。这与南方铁杉幼苗耐荫、小树喜光、生长缓慢的生物学特性分不开[9]。
4 结论与讨论
4.1 南方铁杉群落分类
第四纪全球普遍降温、冰期和间冰期的出现引起了植物群的南北迁移,呈东北-西南走向的武夷山脉因仅受到山岳冰川的微弱影响,而成为南北区系成分来回迁移的天然“桥梁”和重要集散地,从而保存了许多第三纪植物区系的残遗或后裔,有些还有原始林遗留至今[30]。本文在全面群落调查发现30个样地(18 000 m2)共调查到维管植物210 种,隶属于68 科112 属,其中不乏有南方红豆杉、天目紫茎、等珍稀植物,也有柳杉、大血藤等古老孑遗植物。同时,采用TWINSPAN 分类与DCCA 排序方法,将30个南方铁杉群落划分成11 个群落类型。说明在漫长地质历史进程中,南方铁杉与许多物种共同进化,形成多种多样的群落类型。
另外还发现海拔、坡位等环境因子与土壤养分、厚度等土壤环境因子对群落的形成和分布都有一定的影响,但土壤因子的影响力大于空间因子,前解释能力为45.38%,而后仅为23.13%,二者相耦合作用占7.65 %。这充分反映了在山体尺度上,海拔、坡位、坡度、坡向等对光、热、水和土壤养分等资源因子的再分配,影响植被类型的因子大大增多[31],在陡坡、崖锥和沟谷等微地形单元一般有频繁的滑坡、坡塌和侵蚀等干扰[32],这些干扰限制了很多地带性顶极物种的定居,并且弱化了物种的竞争能力,南方铁杉因阳性、长寿和菌根丰富等原因,可在此长期生存,形成特殊的顶极群落。
4.2 南方铁杉种群更新能力评价与分析
更新能力是衡量种群延续能力的重要指标,是珍稀植物保护生态学的重要内容[33]。多数木本植物可通过种子、桩萌、根萌等方式实现种群更新[15],但南方铁杉萌蘖能力差,更新过程全靠种子萌发及幼苗的健康成长[9]。因此对其种萌发与幼苗生长有影响的环境因子都会影响到南方铁杉的更新。
本研究发现南方铁杉更新能力与群落所在的空间位置、土壤养分状况关系不明显,而与所在群落的组成与结构密切相关。乔木层、灌木层落叶树优势度越大、郁闭度越低,南方铁杉更新能力越强;反之,常绿树优势度越大、林下郁闭度越高,南方铁杉更新能力就越弱。所以在枫香、光皮桦、山樱花、雷公鹅耳枥等占优势的群落类型(类型II)中南方铁杉更新能力最强;在乔木层常绿树种占优势,而灌木层为落叶树种(吊钟花、江南山柳、满山红等)的群落类型(类型Ⅸ),南方铁杉更新也较正常;但乔木层、灌木层优势种都是常绿树种的群落(类型Ⅲ、Ⅶ、Ⅷ)中,南方铁杉更新能力较弱,尤其是林下受毛秆箭竹、箬竹入侵时(群落类型Ⅳ、Ⅴ、Ⅵ、Ⅷ、Ⅺ),林下幼苗几乎没有,种群更新能力降到最低,几乎不能正常更新。这一方面与南方铁杉阳性喜光、生长缓慢生物学习性有关,另一方面也可能与竹类植物竞争力、凋落物性质等有关[34-38]。因此,南方铁杉更新能力与群落组成与结构有密切关系。
同时,本研究回答了南方铁杉更新差异的问题。认为南方铁杉林更新困难[6,11-12]是因为他们在群落取样时,只取发育盛期或后期的南方铁杉群落,因常绿阔叶树占优势,或者受到竹类植物入侵[11]。所以南方铁杉更新困难。如果取样时群落类型比较多[3]或调查范围足够宽,则发现南方铁杉能正常更新[21]。
4.3 保护措施与建议
保护生物学理论认为,Meta 种群的灭绝通常是从局部种群的灭绝开始的,各局域种群的稳定性都关乎整个物种的延续[39]。大面积南方铁杉不仅是本区特殊森林景观,也为研究上亿年来南方铁杉及其伴生种的共同演化提供了珍贵的研究素材。因此,从群落水平上强化对残存南方铁杉既是保护南方铁杉种群的需要,也是保护其它珍稀植物及复杂种关系必需。
虽然南方铁杉存在结实率不高、种子生活力低等,造成幼苗幼树分布少[40],但部分群落却正常更新,显然南方铁杉自身的生物学特性,并不足以构成其濒危的主要原因。根据分类和更新能力分析,南方铁杉具有典型阳生树种的特征,群落结构及其光资源分配,可能是自然更新的重要限制因子。本文认为群落结构调整、增加透光度是促进南方铁杉更新的关键。对早期以落叶阔叶树为主的群落,因其群落透光度较强,不需采用辅助措施,南方铁杉就会自然更新、良好生长;对常绿阔叶树较多的群落,因其郁闭度较大、透光度低,应适当进行适度的人为措施(择伐、间伐)部分猴头杜鹃、鹿角杜鹃、薄叶山矾、皖闽八角、多脉青冈、石栎、格药柃等常绿阔叶树,以降低林地郁闭度,改善群落通风条件,促进其种子传播,促使铁杉种子萌发;对现在遭受毛秆箭竹、箬竹入侵的群落,应将之伐除,对将面临入侵干扰的群落并采取措施防止竹类植物扩张,伐除林外竹子[41]。因此,在迁地保护南方铁杉群落效果有限的情况下,如何模拟自然,创造微生境条件[42],促进南方铁杉种群的实生苗更新是一个亟待研究的课题。
[1]李林初.若干铁杉属植物核型的比较研究[J].广西植物,1988,8(4):324-328.
[2]郑万钧.中国树木志[M].北京:中国林业出版社,1983.
[3]张志祥,刘鹏,蔡妙珍,等.九龙山珍稀濒危植物南方铁杉种群数量动态[J].植物生态学报,2008,32(5):1146-1156.
[4]封磊,洪伟,吴承祯,等.南方铁杉种群结构动态与空间分布格局[J].福建林学院学报,2008,28(2):110-114.
[5]罗金旺.福建光泽南方铁杉天然林的生长规律与生物量[J].福建林学院学报,2011,31(2):156-160.
[6]黄宪刚,谢强.猫儿山南方铁杉种群结构和动态的初步研究[J].广西师范大学学报:自然科学版,2000,18(2):86-90.
[7]冯祥麟,胡刚,刘正华.贵阳高坡南方铁杉群落特征及种群动态调查研究[J].贵州林业科技,2011,39(2):26-29.
[8]钱晓鸣,黄耀坚,张艳辉,等.武夷山自然保护区南方铁杉外生菌根生物多样性[J].福建农林大学学报:自然科学版,2007,36(2):180-185.
[9]李晓铁.猫儿山林区南方铁杉生态学特性初步调查研究[J].广西林业科技,1990(4):22-23,19.
[10]李晓铁,玉伟朝,罗远周,等.南方铁杉扦插繁殖技术[J].林业实用技术,2008(6):21-22.
[11]郭连金,洪森荣,夏华炎.武夷山自然保护区濒危植物南方铁杉种群数量动态分析[J].上饶师范学院学报:自然科学版,2006(6):74-78.
[12]杜道林,刘玉成,刘川华.茂兰喀斯特山地南方铁杉种群结构和动态初探[J].西南师范大学学报:自然科学版,1994,19(2):169-174.
[13]Hanson H C,Churchill E D.The Plant Community[M].New york:Reinhold Publishing Corporation,1961.
[14]Neuhauser C,Andow D A.George E H,et al.Community genetics:expanding the synthesis of ecology and genetics[J].Ecology,2003,84(3):545-558.
[15]李小双,彭明春,党承林.植物自然更新研究进展[J].生态学杂志,2007,26(12):2081-2088.
[16]刘万德,李帅锋,张志钧,等.滇西北云南红豆杉群落结构与更新特征[J].生态学杂志,2012(12):3024-3031.
[17]杨永川,穆建平,杨轲.残存银杏群落的结构及种群更新特征[J].生态学报,2011,31(21):6396-6409.
[18]张志祥,刘鹏,刘春生,等.浙江九龙山南方铁杉(Tsuga tchekiangensis)群落结构及优势种群更新类型[J].生态学报,2008,28(9):4547-4558.
[19]杨跃军,孙向阳,王保平.森林土壤种子库与天然更新[J].应用生态学报,2001,12(2):304-308.
[20]王传华,李俊清,陈芳清,等.鄂东南低丘地区枫香林下枫香幼苗更新限制因子[J].植物生态学报,2011,35(2):187-194.
[21]袁荣斌,邹思成,兰文军,等.江西武夷山国家级自然保护区南方铁杉资源调查初报[J].江西林业科技,2012,39(4):37-39.
[22]Michele D S,Achmed A,Alessio F,et al.Classification and distribution patterns of plant communities on Socotra Island,Yemen[J].Applied Vegetation Science,2013,16(1):148-165.
[23]Pajunen A M,Kaarlejärvi E M,Forbes B C,et al.Compositional differentiation,vegetation-environment relationships and classification of willow-characterised vegetation in the western Eurasian Arctic[J].Journal of Vegetation Science,2010,21(1):107-119.
[24]鲁如坤.土壤农业化学分析方法.[M],北京:中国农业科技出版社,2000.
[25]王伯荪,余世孝,彭少麟,等.植物群落学实验手册[M].广州:广东高等教育出版社,1996.
[26]Leps J,Smilauer P.Multivariate analysis of ecological data using CANOCO[M].New York:Cambridge University Press,2003.
[27]Ahmad S S.Ordination and classification of herbaceous vegetation in Margalla Hills National Park Islamabad Pakistan[J].Biological Diversity and Conservation,2009,2(2):38-44.
[28]邓贤兰,杨 吴,刘玉成,等.江西野生种子植物区系多样性及其基本特征[J].植物科学学报,2012,30(1):22-30.
[29]杜丽侠,邢韶华.北京八达岭地区灌木群落空间分布格局与土壤环境因子的关系分析[J].西北植物学报,2009,29(3):601-607.
[30]江西森林编委会.江西森林[M].北京:中国林业出版社;南昌:江西科学技术出版社,1986.
[31]Clark D B,Palmer M W,Clark D A.Edaphic factors and the landscape-scale distributions of tropical rain forest trees[J].Ecology,1999,80(8):2662-2675.
[32]区余端,苏志尧,李镇魁,等.地形因子对粤北山地森林不同生长型地表植物分布格局的影响[J].应用生态学报,2011,22(5):1107-1113.
[33]李玲,张光富,锐王,等.天目山自然保护区银杏天然种群生命表[J].生态学杂志,2011,30(1):53-58.
[34]Tao J P,Shi X P,Wang Y J.Effects of different bamboo densities on understory species diversity and trees regeneration in an Abies faxoniana forest,Southwest China[J].Scientific Research and Essays,2012,7(6):660-668.
[35]Caccia F D,Chaneton E J,Kitzberger T.Direct and indirect effects of understorey bamboo shape tree regeneration niches in a mixed temperate forest[J].Oecologia,2009,161(4):771-780.
[36]Wang Y J,Shi X P,Peng Y,et al.Effects of fine-scale pattern of dwarf bamboo on understory species diversity in Abies faxoniana Forest,Southwest,China[J].Sains Malaysiana,2012,41(6):649-657.
[37]Lima R A F,Rother D C,Ana E M,et al.Bamboo overabundance alters forest structure and dynamics in the Atlantic Forest hotspot[J].Biological Conservation,2012,147(1):32-39.
[38]刘晋仙,郭庆学,王玉平,等.竹类凋落叶对4 种乔木树种种子萌发的影响[J].林业科学,2013,49(1):33-40.
[39]赵淑清,方精云,雷光春.物种保护的理论基础:从岛屿生物地理学理论到集合种群理论[J].生态学报,2001,21(7):1171-1179.
[40]李晓铁.猫儿山林区南方铁杉生长调查初报[J].广西林业科技,1992,21(1):24-26.
[41]Kisanuki H,Kudo T,Nakai A.Removing aboveground vegetation facilitates survival but slows height growth of spruce saplings in a fenced,degraded sub-alpine forest in central Japan[J].Journal of Forest Research,2012,17(1):110-115.
[42]Tang C Q,Ohsawa M.Tertiary relic deciduous forests on a humid subtropical mountain,Mt.Emei,Sichuan,China[J].Folia Geobotanica,2002,37(1):93-106.
猜你喜欢
建筑与预算(2022年6期)2022-07-08
现代园艺(2021年23期)2021-12-01
林业勘查设计(2020年1期)2021-01-18
新农业(2020年18期)2021-01-07
农村实用技术(2020年1期)2020-12-16
乡村科技(2018年8期)2018-02-13
诗潮(2017年2期)2017-03-16
湖南林业科技(2017年1期)2017-02-06
河南科技(2014年16期)2014-02-27
河池学院学报(2014年5期)2014-02-27
