
大肠杆菌TorS/TorR二组分体应答蛋白TorR对DNA复制起始的影响
2015-02-04 06:12姚远乔佳鑫李静李慧莫日根
遗传 2015年3期
姚远,乔佳鑫,李静,李慧,莫日根
大肠杆菌TorS/TorR二组分体应答蛋白TorR对DNA复制起始的影响
姚远,乔佳鑫,李静,李慧,莫日根
内蒙古大学生命科学学院,呼和浩特 010021
二组分体作为一种信号转导系统在细菌中普遍存在,能够感知外界环境变化并做出应答。细菌中CckA/CtrA、ArcA/ArcB和PhoP/PhoQ二组分体与DNA复制起始和细胞分裂相关,但目前还未见TorS/TorR二组分体对细胞周期及DNA复制影响的相关报道。大肠杆菌TorS/TorR二组分体能够监测细胞周围氧化三甲胺(Trimethylamine oxide, TMAO)的浓度变化,但其是否影响DNA复制起始呢?文章利用流式细胞仪检测了Δ和Δ突变体菌株的复制式样。结果发现,Δ突变菌株每个细胞复制起始原点数目和倍增时间与野生型细胞一致,而Δ突变菌株每个细胞复制起始原点数目多于野生型细胞,说明复制起始发生时间比野生型细胞早。但是过表达TorR蛋白或者共同表达TorS和TorR蛋白都不能使Δ突变体表型恢复为野生型表型。而在野生型和Δ突变细胞中过表达SufD蛋白能使复制起始提早发生,在Δ和Δ双突变细胞中复制起始延迟。所以,TorR可能通过改变基因的表达来间接影响染色体复制起始。
TorS/TorR二组分体;应答蛋白TorR;DNA复制;大肠杆菌
细菌时时刻刻都被各种分子所包围,这些分子或者来自于周围环境,或者来自于细菌本身的代谢产物。所以,细菌要想保证自身存活和繁殖,就必须做出应答来适应周围不断变化的环境。而这些应答途径的激活大多都是先接受一个外界信号分子的刺激,然后激活特异性基因的转录。在细菌中,由结合于细胞膜的组氨酸激酶(Histidine kinase)和其相互协作的应答蛋白(Response regulator)组成的二组分体(Two-component system)可以感知温度、渗透压和pH值等环境变化,从而激活(或抑制)靶基因表达,使细胞做出相应的应答[1]。二组分体信号转导通过以下步骤来实现:接受外界信号刺激的组氨酸激酶将ATP上磷酸基团转移到其特定的组氨酸残基上,相继磷酸基团会被转移到应答蛋白的天冬氨酸残基,磷酸化会引起应答蛋白构型的改变,从而影响其与靶基因启动子结合的能力[2],由此诱导细胞对环境变化做出反应。
大肠杆菌TorS/TorR二组分体中TorS是一个跨膜的组氨酸蛋白激酶,TorS通过N-末端特异性感知细胞外环境中的氧化三甲胺(Trimethylamine oxide, TMAO)浓度,并使自身His443位点磷酸化,依次将磷酸基团传递到自身Asp723和C-末端的His850位点,最终转移到TorR接收区域的Asp53位点。磷酸化的TorR蛋白可以诱导操纵子的表达,从而应答细胞周围TMAO的浓度变化[3]。
研究表明,细菌中CckA/CtrA、ArcA/ArcB和PhoP/PhoQ二组分体与DNA复制起始和细胞分裂相关[4~6]。但目前还未见TorS/TorR二组分体对细胞周期及DNA复制影响的相关报道。本文发现,相对野生型细胞,Δ突变细胞的复制起始提早发生,但在Δ突变细胞中,过量表达TorR不能使Δ突变表型恢复。进一步研究表明,TorR可能通过调节基因的表达量来间接影响DNA复制起始。
1 材料和方法
1.1 菌株与质粒
本研究使用的所有菌株均为K-12。野生型菌株BW25113和其派生的::R、::R、::R、::R、::R、::R、::R、::R、::R、::R缺失突变菌株由美国亚利桑那州立大学施一燊博士惠赠。pACYC177[7]和pcDNA3-mCherry[8]质粒由本实验室保存,p、p[9]和pUHE21- 2lac[10]质粒由施一燊博士惠赠。
1.2 方法
1.2.1 质粒构建
将基因连同其自身的启动子利用一对特异性引物(正向:5¢-CCGCTCGAGTTGCTCGCTTCCAGTTTG-3¢;反向:5¢-CGGGATCCTGTCAGCCCACCGATTTT-3¢),以BW25113基因组DNA为模板进行PCR扩增,扩增得到的PCR产物经Ⅰ和HⅠ限制性内切酶消化后连接到同样用这两种酶消化后的pACYC177片段上,获得p过表达质粒(图1A)。为了构建TorR-mCherry融合表达质粒,先将基因连其自身启动子同时缺失终止密码子的片段利用特异性引物(正向:5¢-CCCAAGCTTGCGCCAGTACCGACCAACG-3¢;反向:5¢-CGGGGTACCGCACACATCAGCGGCTAAG-3¢)以BW25113菌株基因组DNA为模板经过PCR扩增,得到两端分别带有dⅢ和Ⅰ酶切位点的片段。然后,利用特异性引物(正向:5¢-CGGGGTACCGTGAGCAAGGGCGAGGA-3¢;反向:5¢-CGGGATCCTTACTTGTACAGCTCGTCCATGC-3¢)以pcDNA3-mCherry质粒为模板经过PCR扩增,得到两端分别带有Ⅰ和HⅠ酶切位点的片段。将上述两条片段经过相应限制性内切酶的消化、连接,获得-片段,之后将其插入到pACYC177质粒的dⅢ和HⅠ限制性内切酶位点间,构建了融合表达质粒p-mCherry(图1B)。根据基因的开放阅读框(ORF)序列设计引物(正向:5¢-CGGGATCCGTGAGTAAACGTTATTTTGTCACCGG-3¢;反向:5¢-CCCAAGCTTCTACAACAAGGCAAGGTTTATGTAC-3¢),以BW25113基因组DNA为模板进行PCR扩增,随后用HⅠ和dⅢ限制性内切酶消化,并连接到pUHE21-2lac质粒启动子序列之后,获得p质粒,的表达需要IPTG诱导(图1C)。p和p质粒来源于ASKA库,其或由IPTG诱导表达。
构建好的质粒由上海生工生物工程技术服务有限公司进行测序。利用NCBI(www.ncbi.nim.nih.gov/)在线分析软件进行序列比对,发现插入的基因未发生突变,质粒构建正确。
1.2.2 双突变菌株的构建
先将Δ突变菌株的卡那霉素抗性删除[11],之后利用P1转导构建得到ΔΔ双突变菌株[12],本文共得到23个双突变菌株,随机选取4个单菌落,利用PCR和流式细胞仪检测方法对其进行验证,证明所得到的双突变菌株构建正确。
1.2.3 细胞倍增时间(Doubling time)的测定
将过夜培养的菌液按照1:10000倍数在ABTGcasa培养基[13]中稀释,然后转入37℃水浴摇床中培养。每隔15 min检测活体细胞数目,用紫外分光光度计(Shimadzu, UV-1800)测量值(ABTGcasa培养基450),记录相应的时间。如果质粒需要IPTG诱导表达,在450≈0.03时加入终浓度为0.1 mmol/L的IPTG。450≈0.04开始,450≈0.5结束。测得的值取底数为2的log值,与时间差(min)分别为横坐标和纵坐标,用Microsoft EXCEL作图得到细胞生长曲线,线性方程的斜率即为细胞倍增时间。重复3次,得到的平均值为菌株在ABTGcasa培养基上的细胞倍增时间,比较野生型与突变体的细胞倍增时间。
1.2.4 流式细胞仪测定复制式样和相对荧光强度
在ABTGcasa培养基中37℃培养细胞至对数生长期,在450≈0.15时加入利福平(300 μg/mL)和先锋霉素(10 μg/mL),继续让细胞生长3~4代。利福平能够抑制新的复制起始,但允许已经开始的复制继续进行,先锋霉素能够阻止细胞分裂。收集细胞用70%乙醇固定,用Tris-HCl buffer(pH7.5)洗涤细胞,之后用Hoechst 33258 DNA染料染色30 min,用流式细胞仪(BD LSRFortessa)测定DNA复制式样,具体方法和原理见文献[14]。检测相对荧光强度的样品与测定复制式样的样品相同,利用流式细胞仪PE-Texas Red通道检测mCherry的相对荧光强度。
1.2.5 荧光显微镜样品的制备及观察
荧光显微镜样品与流式细胞仪检测样品的处理与制备方法相同,取10 μL固定样品制片,利用荧光显微镜(蔡司Axio Imager A2)观察,滤光片43(chroma filter sets 43,Carl Zeiss)观察mCherry。同时用AxioCam MRc5相机拍照。最后使用AxioVision Rel. 4.8软件进行数据分析和图片处理。
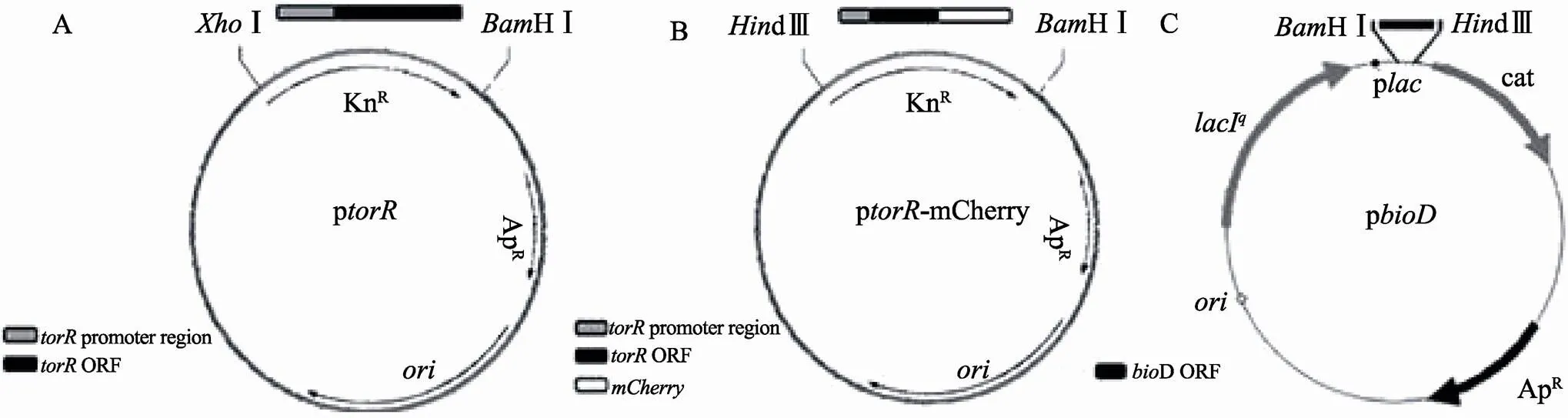
图1 过表达质粒构建
A:p质粒构建。图中黑色长方形代表基因的开放阅读框序列,灰色长方形代表基因启动子。利用PCR扩增出基因及其自身启动子的片段经过Ⅰ和HⅠ消化连接到pACYC177质粒上,经过改造的质粒具有氨苄青霉素抗性基因。B:p-mCherry质粒构建。图中黑色长方形代表基因的开放阅读框序列(ORF),灰色长方形代表基因启动子,白色长方形代表基因,经过改造的质粒具有氨苄青霉素抗性基因。C:p质粒构建。图中黑色长方形代表基因的ORF区域,插入到pUHE21-2lac质粒中HⅠ和dⅢ酶切位点之间,并位于启动子(p)之后,具有氨苄青霉素和氯霉素抗性基因。
2 结果与分析
2.1 ΔtorR和ΔtorS突变菌株的鉴定
为了鉴定Δ和Δ突变菌株的正确性,本文分别以和基因的ORF序列为模板在基因内部设计特异性引物,扩增大小合适的片段来检测相应基因是否被敲除。利用特异性引物(正向:5¢-GGATTGACCGTATTGTTGG-3¢;反向:5¢-GGTGGCATTTGTGACGAA-3¢),分别以BW25113和Δ菌株基因组DNA为模板,扩增基因内部262 bp片段(图2A)。利用特异性引物(正向:5¢-TAATGAGCGGACGGGTGA-3¢;反向:5¢-TGGGTTAGCGGGTTATCTT-3¢),分别以BW25113和Δ菌株基因组DNA为模板,扩增基因内部490 bp片段(图2B)。如图2A所示,利用野生型BW25113基因组DNA(阳性对照)为模板进行目的基因的扩增,有明显的目的条带,以ddH2O(阴性对照)为模板进行扩增,未见到目的条带,利用Δ菌株基因组DNA为模板进行目的片段扩增,也未检测到目的条带。图2B以Δ菌株基因组DNA为模板进行目的基因的扩增,并未检测到目的条带。以上结果证明,本文所获得的缺失突变菌株构建正确。
2.2 torR基因的缺失导致DNA复制起始的提早发生
为了检测TorR对DNA复制起始的影响,将野生型、Δ和Δ细胞在ABTGcasa培养基中培养到对数期,加入利福平(300 μg/mL)和先锋霉素(10 μg/mL)培养细胞3~4代。经Hoechst 33258 DNA染料染色后,用流式细胞仪检测野生型、Δ和Δ突变细胞的复制式样。结果,发现野生型菌株主要有2、4、8条染色体的细胞,而在Δ突变体中只发现4、8条染色体的细胞,而且其细胞倍增时间(30 min)比野生型细胞(32 min)短,说明TorR缺失导致复制起始的提早发生[14](图3)。但是,Δ突变菌株和野生型细胞的复制式样基本相同(图3),其细胞倍增时间和野生型细胞同为32 min,显然,TorS的缺失并不改变DNA复制起始和细胞倍增时间。这些结果说明,Δ突变菌株复制起始提早发生不是因为TorS/TorR二组分体信号转导途径阻断所引起。
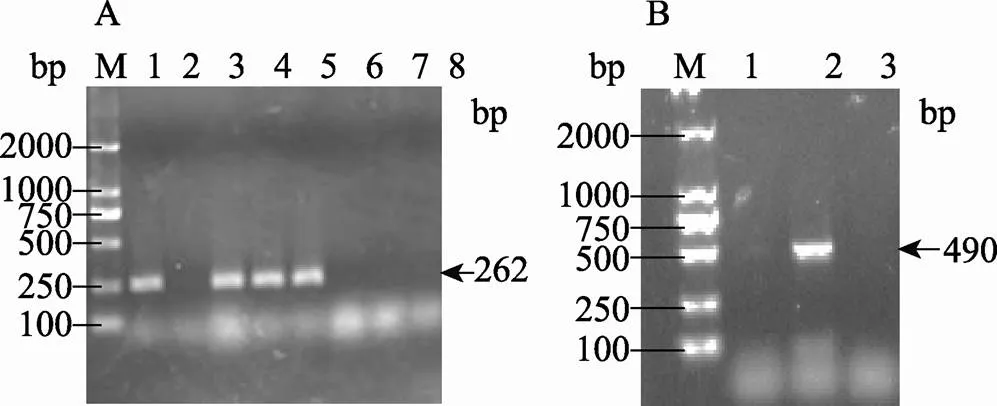
图2 突变菌株的鉴定
A:Δ突变菌株的鉴定。泳道1、3~5:利用野生型BW25113基因组DNA(阳性对照)为模板进行目的基因的扩增;泳道2:阴性对照;泳道6~8:利用Δ菌株基因组DNA为模板进行目的片段扩增。B:Δ突变菌株的鉴定。泳道1:阴性对照;泳道2:阳性对照,以野生型BW25113基因组DNA为模板进行目的片段的扩增;泳道3:以Δ菌株基因组DNA为模板进行目的基因的扩增。
2.3 质粒表达的TorR蛋白不能将ΔtorR突变表型恢复为野生型表型
将pACYC177和p过表达质粒利用CaCl2法分别转化到Δ突变体和野生型细胞中,按图3中所述方法,用流式细胞仪测定两者的染色体复制式样(图4)。结果表明,pACYC177不会影响野生型和Δ突变菌株的复制式样(图4:A,B),在野生型和Δ突变菌株中过量表达TorR也不能改变其DNA复制式样(图4:C,D)。
2.4 TorR-mCherry融合蛋白的表达也不能恢复ΔtorR突变表型
基于用p质粒转化不能使Δ突变细胞改变其复制起始提早发生的表型,本文推测p质粒可能不表达TorR蛋白。因此,本文构建了能够表达TorR-mCherry融合蛋白的表达质粒p-mCherry,-基因的表达受启动子的控制。为了验证p质粒表达TorR蛋白,而p-mCherry表达TorR-mCherry融合蛋白,首先将p-mCherry质粒利用CaCl2转化法引入Δ突变菌株中,随后用流式细胞仪和荧光显微镜检测mCherry的表达情况(图5A)、表达量(图5C)及Δ/p-mCherry菌株的复制式样(图5B)。如图5A所示,细胞有明显的红色荧光,说明TorR-mCherry蛋白在细胞中有表达;流式细胞仪PE-Texas Red荧光通道也检测到了mCherry发出的特异性荧光信号(图5C),而在对照质粒p中未检测到任何信号。这些结果说明p和p-mCherry质粒分别表达有TorR蛋白和TorR- mCherry融合蛋白。然而,由p-mCherry质粒表达的TorR-mCherry融合蛋白依然不能使Δ突变的提早复制起始表型恢复为野生型表型(图5B)。
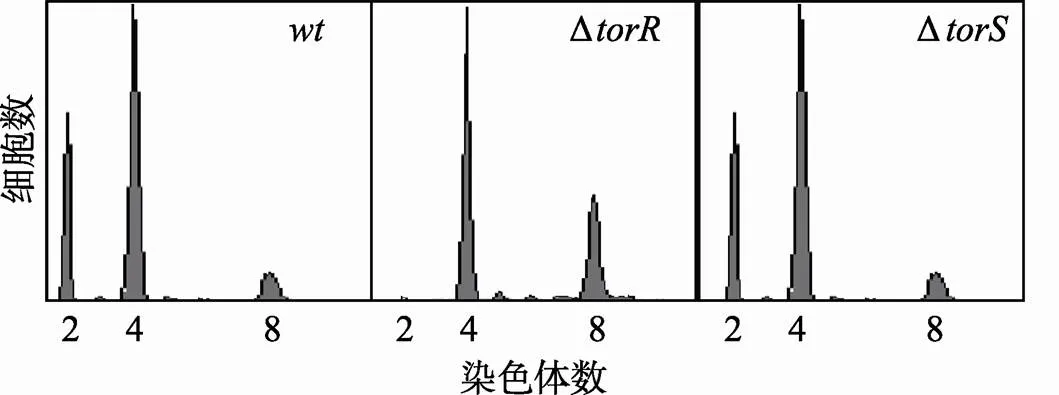
图3 ΔtorR缺失突变引起DNA复制起始提早发生
处在对数生长期的细胞(ABTGcasa培养基)中加入利福平和先锋霉素继续培养3~4代后用70%酒精固定细胞。Hoechst 33258进行染色后,用流式细胞仪对细胞所含的染色体数目进行测定。横坐标表示每个细胞所含染色体数目,纵坐标表示细胞数目。每个测定包含了10 000个细胞。
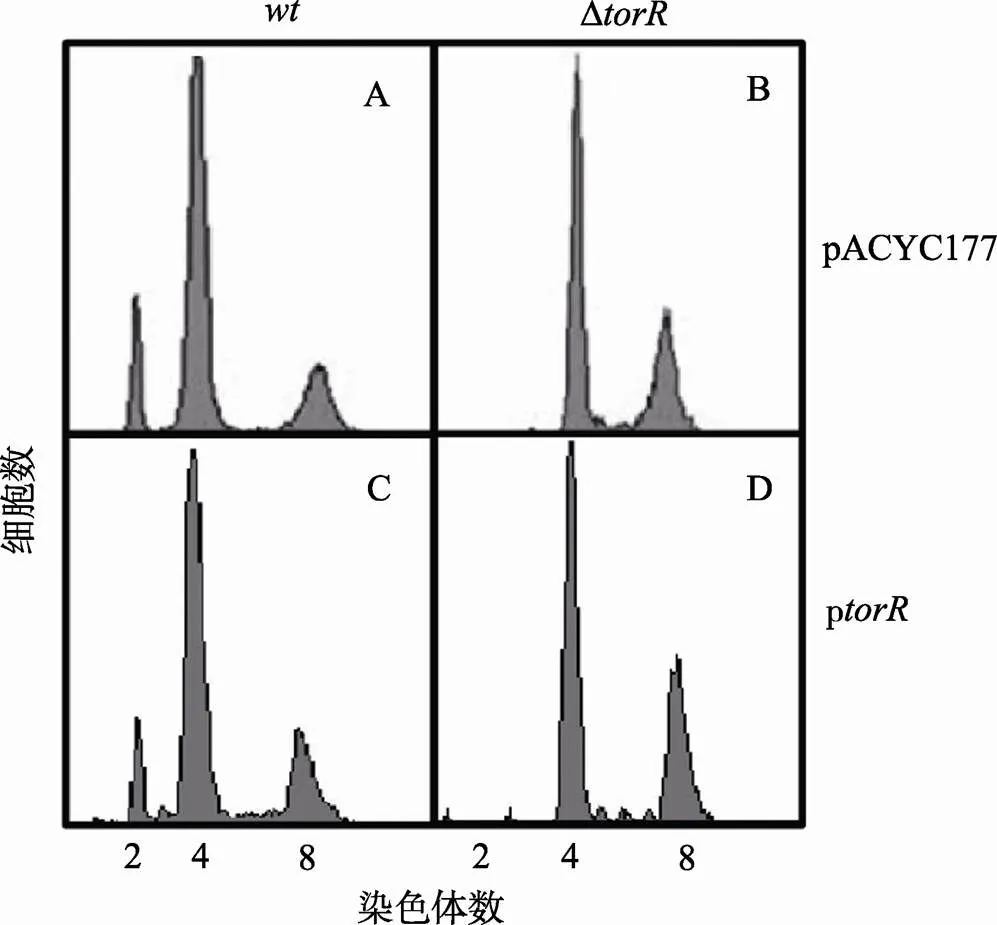
图4 过量表达TorR蛋白不能恢复ΔtorR突变表型
样品处理和测定方法如图3所述,横坐标表示每个细胞所含染色体数目,纵坐标表示细胞数目。A,B:野生型菌株或者Δ突变菌株中转入空质粒pACYC177并不会影响复制式样。C,D:野生型菌株或者Δ突变菌株中转入p质粒,过表达TorR不能影响其复制式样。每个测定包含了10 000个细胞。
2.5 共表达TorR和TorS蛋白也不能使ΔtorR突变表型恢复为野生型表型
为什么过表达TorR不能恢复Δ突变表型?是否由于细胞内TorS含量不足而不能有效磷酸化TorR,进而导致该二组分信号转导通路受阻?为了探讨以上问题,本文将质粒p和p共转化到Δ突变菌株中,用流式细胞仪检测其DNA复制式样。结果发现,同时过表达TorR和TorS蛋白不能使其DNA复制样式改变,即与Δ突变细胞一样,共表达TorR和TorS蛋白的Δ菌株仍然以含4、8条染色体的细胞为主(图6),其原因仍不清楚。
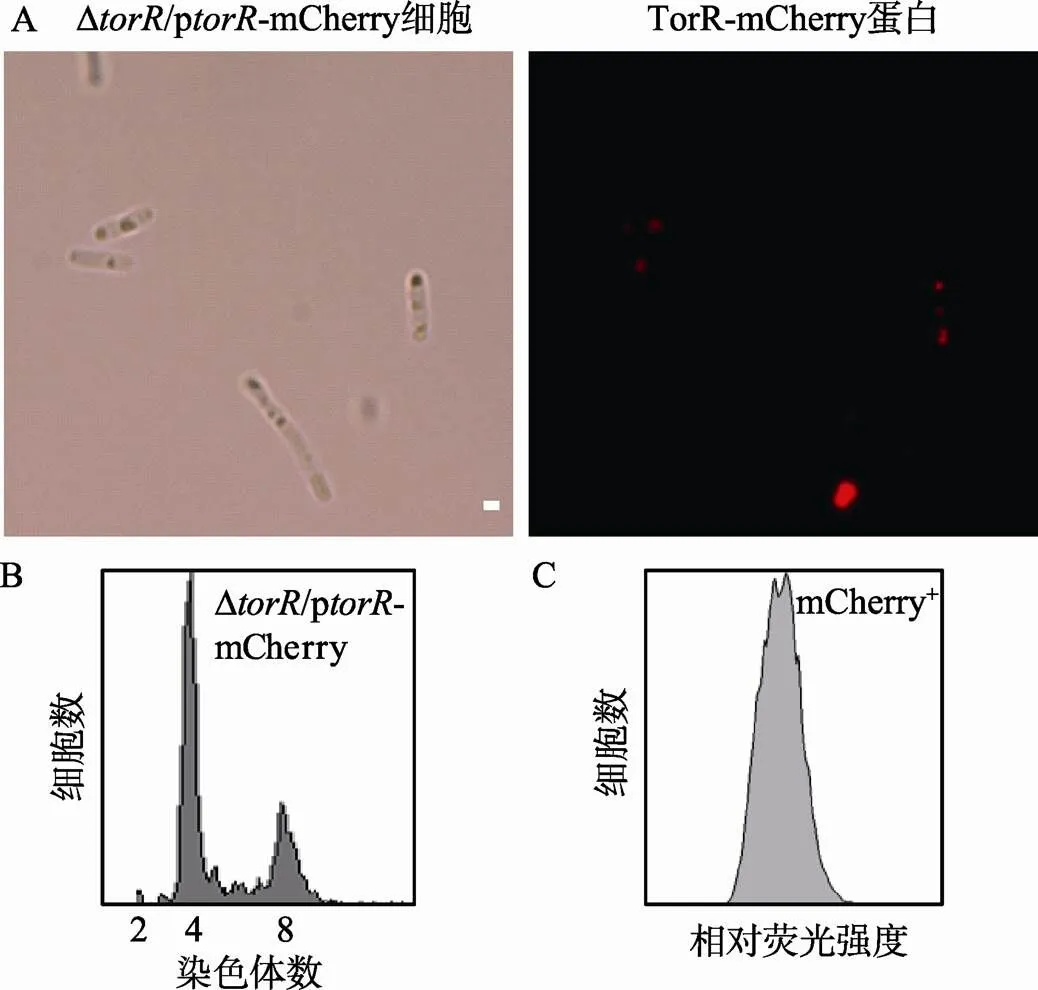
图5 质粒表达的TorR-mCherry融合蛋白不能恢复ΔtorR突变表型
A:荧光显微镜下观察TorR-mCherry融合蛋白的亚细胞定位。图中标尺为1 µm,红色表示TorR-mCherry。B:Δ突变菌株过量表达TorR-mCherry蛋白的DNA复制式样,样品处理方法如图3所述。横坐标表示每个细胞所含染色体数目,纵坐标表示细胞数目。每个测定包含10 000个细胞。C:利用流式细胞仪测定Δ/p-mCherry菌株中TorR-mCherry的相对荧光强度值。横坐标表示mCherry相对荧光强度,纵坐标表示所测定的细胞数目。
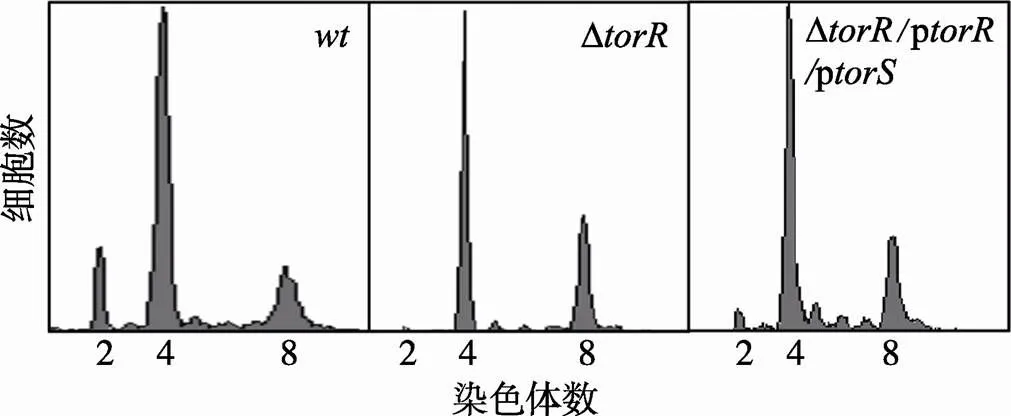
图6 共同表达TorR和TorS蛋白不能恢复ΔtorR突变的表型
样品处理和测定方法如图3中描述,横坐标表示每个细胞所含染色体数目,纵坐标表示细胞数目。:野生型细胞复制式样;Δ:Δ突变细胞的复制式样;Δ/p/p:Δ突变细胞共表达TorR和TorS蛋白后的DNA复制式样。p质粒IPTG诱导浓度为0.1 mmol/L,每个测定包含10 000个细胞。
2.6 sufD和bioD基因缺失导致DNA复制起始滞后
研究表明,基因的缺失影响多个基因的表达,包括、、、、、、和等[15]。所以,TorR蛋白可能通过调节这些基因的表达来间接影响DNA复制起始。为了验证这个推测,本文利用流式细胞仪分别检测了Δ、Δ、Δ、Δ、Δ、Δ、Δ和Δ突变细胞的DNA复制式样。如图7所示,除了Δ和Δ缺失突变体的复制式样不同于野生型细胞外,其余突变菌株的复制式样与野生型并无明显差异。Δ和Δ缺失突变体与野生型相比,缺少含8条染色体的细胞,这说明和基因的缺失会引起DNA复制起始延迟。
2.7 SufD蛋白过量表达引起DNA复制起始提早发生
在Δ缺失突变菌株中,基因的表达量高于野生型细胞[15],此时DNA复制起始早于野生型细胞。因此,基因表达量的高低可能与DNA复制起始时间有直接联系。本文分别在野生型菌株和Δ缺失突变菌株中过量表达SufD蛋白,用流式细胞仪检测其复制式样。结果发现,相较于野生型细胞,/p和Δ/p菌株DNA复制起始发生的时间都有明显提前(图8)。与/p细胞比较,Δ/p菌株有更多的细胞有8条染色体,说明缺失TorR和过表达SufD的效果是累加的。而过表达BioD并不明显改变Δ缺失突变体细胞的DNA复制式样(图9)。
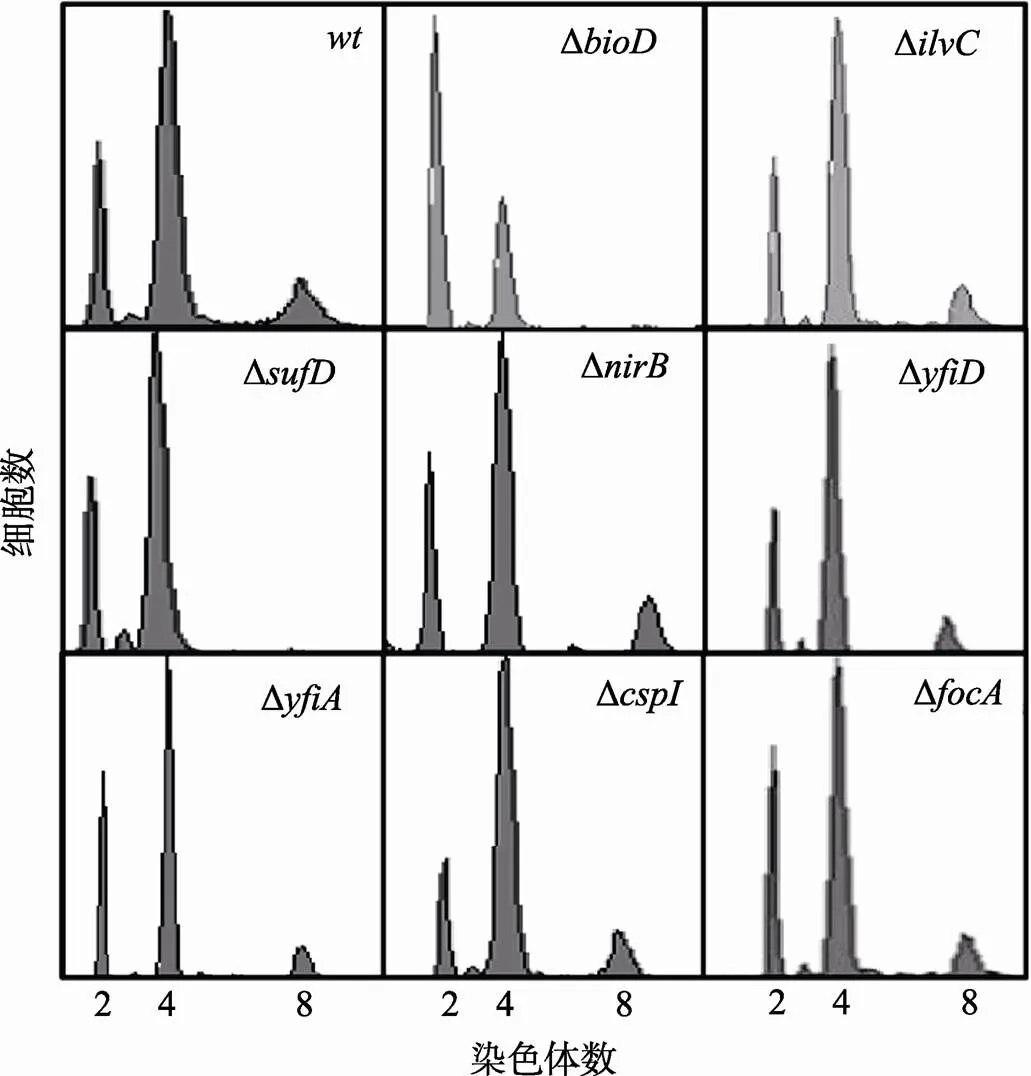
图7 sufD基因缺失导致DNA复制起始延迟
样品处理和测定方法如图3中描述,野生型菌株和突变体菌株在图中已经标出,横坐标表示每个细胞所含染色体数目,纵坐标表示细胞数目。每个测定包含了10 000个细胞。
2.8 ΔtorRΔsufD双突变会引起DNA复制起始延迟
利用P1转导法[12],构建了ΔΔ双突变体,并按图3中所述方法检测复制式样,发现ΔΔ双突变体中只存在含有2、4条染色体的细胞(图10),与野生型细胞相比其DNA复制起始延迟,与Δ单突变细胞的复制式样一致,说明对DNA复制起始而言,SufD蛋白的作用独立于TorR蛋白。以上结果支持TorR蛋白通过调节基因表达间接影响DNA复制起始。
3 讨 论
本研究表明,Δ突变细胞的复制起始相较野生型细胞会提早发生,而Δ突变细胞的复制式样与野生型相同。并且在Δ突变细胞中,不论过量表达TorR,还是共同表达TorR和TorS蛋白,都不能使Δ突变表型恢复。前人研究表明,基因能够进行自我调控,即当细胞中TorR过量时,不论TorR是否被磷酸化,都会结合到基因启动子的上,阻断的转录[16],这或许可以解释过量表达TorR并不能在互补实验中看到表型恢复的结果。SufD蛋白与SufB和SufC蛋白共同形成复合体,参与铁硫簇的组装[17]。当细胞中缺乏SufD蛋白时,会引起复制起始延迟,而SufD过表达则使细胞复制起始提早发生,有研究表明基因缺失会导致基因的表达提高[15]。因此,TorR可能通过调节基因的表达量来影响铁硫簇的组装从而间接调控DNA复制起始。而SufD蛋白或者铁硫簇如何调控DNA复制还需要进一步探究。本文还发现Δ缺失突变导致DNA复制起始延迟。而在细胞中基因表达量低于野生型细胞[15],但其DNA复制起始提早发生,也就是说,基因的低表达并未导致复制起始提早发生。另外,在Δ缺失突变细胞中过表达BioD也未见复制式样的改变,这些结果建议TorR并不通过基因表达来影响DNA复制起始。
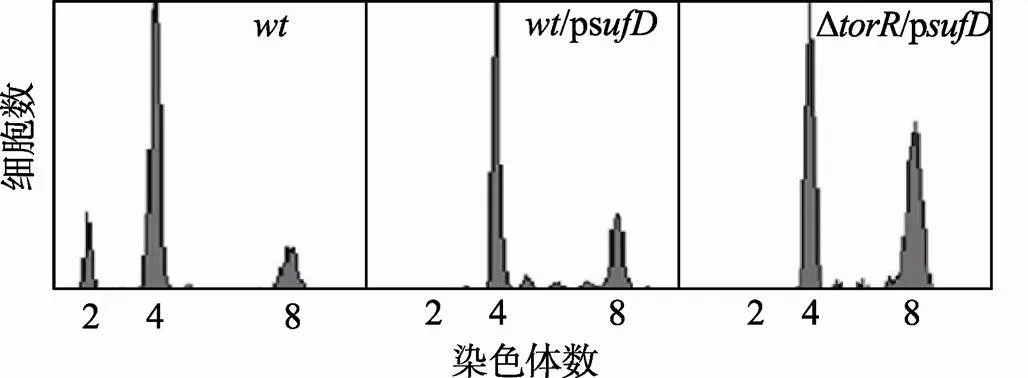
图8 过量表达SufD蛋白引起DNA复制起始提早发生
样品处理和测定方法如图3中描述,野生型菌株,突变体菌株和质粒在图中已经标出,p质粒IPTG诱导浓度为0.1 mmol/L,横坐标表示每个细胞所含染色体数目,纵坐标表示细胞数目。每个测定包含了10 000个细胞。
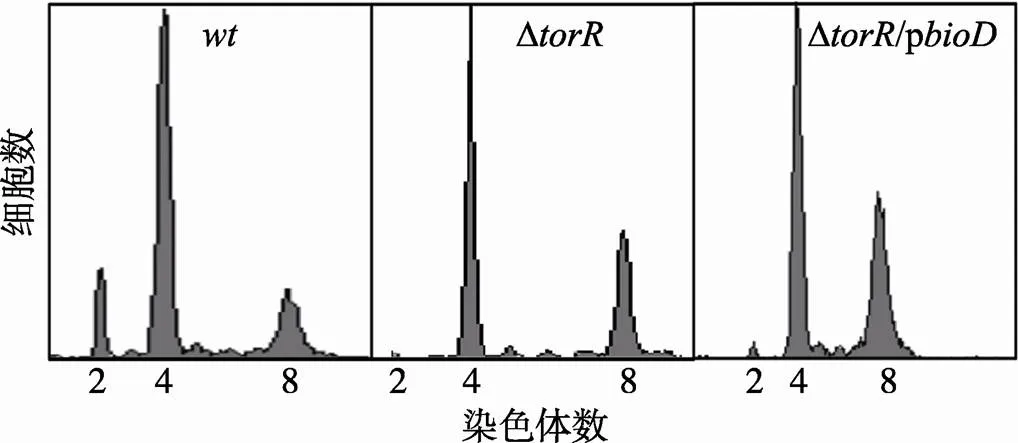
图9 过量表达BioD蛋白不能恢复ΔtorR突变表型
样品处理和测定方法如图3中描述,p质粒IPTG诱导浓度为0.1 mmol/L,横坐标表示每个细胞所含染色体数目,纵坐标表示细胞数目。每个测定包含了10 000个细胞。
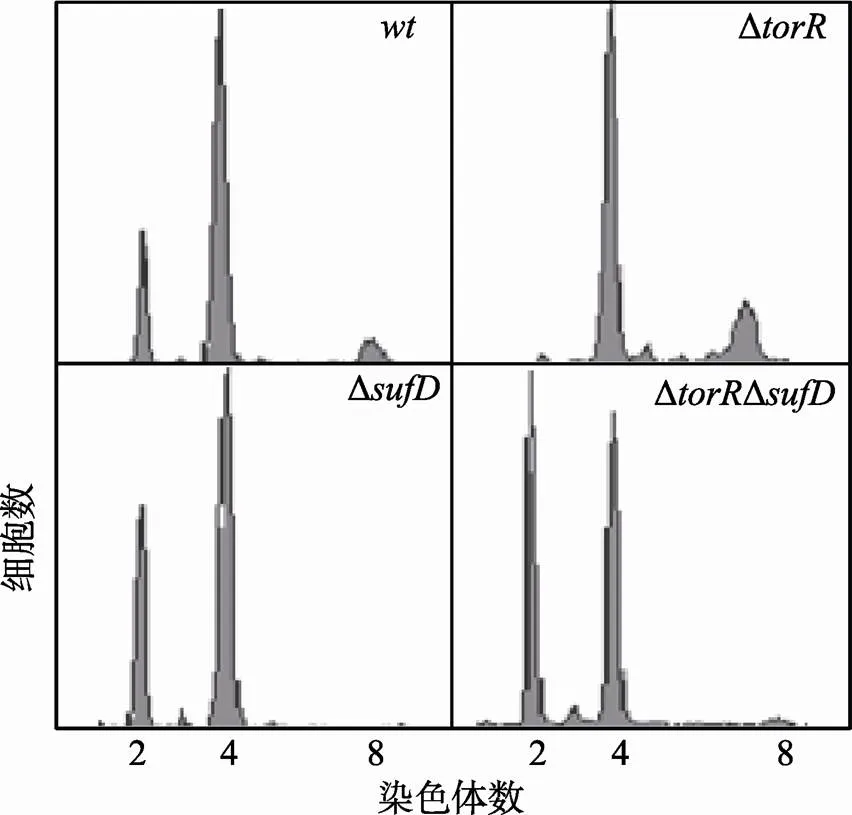
图10 ΔtorRΔsufD双突变会引起DNA复制起始延迟
样品处理和测定方法如图3中描述,横坐标表示每个细胞所含染色体数目,纵坐标表示细胞数目。每个测定包含了10 000个细胞。
[1] Stock AM, Robinson VL, Goudreau PN. Two-component signal transduction., 2000, 69: 183–215.
[2] Buckler DR, Anand GS, Stock AM. Response-regulator phosphorylation and activation: a two-way street?, 2000, 8(4): 153–155.
[3] Jourlin C, Ansaldi M, Méjean V. Transphosphorylation of the torR response regulator requires the three phosphorylation sites of the torS unorthodox sensor in., 1997, 267(4): 770–777.
[4] Jacobs C, Ausmees N, Cordwell SJ, Shapiro L, Laub MT. Functions of the CckA histidine kinase incell cycle control., 2003, 47(5): 1279–1290.
[5] Soncini FC, Groisman EA. Two-component regulatory systems can interact to process multiple environmental signals., 1996, 178(23): 6796–6801.
[6] Colloms SD, Alén C, Sherratt DJ. The ArcA/ArcB two-component regulatory system ofis essential for Xer site-specific recombination at psi., 1998, 28(3): 521–530.
[7] Morigen, Boye E, Skarstad K, Løbner-Olesen A. Regulation of chromosomal replication by DnaA protein availability in: effects of theregion., 2001, 1521(1-3): 73–80.
[8] Zhang XM, Kong W, Ashraf S, Curtiss R Ⅲ. A one-plasmid system to generate influenza virus in cultured chicken cells for potential use in influenza vaccine., 2009, 83(18): 9296–9303.
[9] Kitagawa M, Ara T, Arifuzzaman M, Ioka-Nakamichi T, Inamoto E, Toyonaga H, Mori H. Complete set of ORF clones ofASKA library (a complete set ofK-12 ORF archive): unique resources for biological research., 2006, 12(5): 291–299.
[10] Soncini FC, Véscovi EG, Groisman EA. Transcriptional autoregulation of theoperon., 1995, 177(15): 4364–4371.
[11] Datsenko KA, Wanner BL. One-step inactivation of chromosomal genes inK-12 using PCR products., 2000, 97(12): 6640–6645.
[12] Miller RV, Ripp S, Replicon J, Ogunseitan O, Kokjohn TA. Virus-mediated gene transfer in freshwater environments. In: Gauthier MJ, ed. Gene Transfers and Environment. New York: Springer, 1992: 51–62.
[13] Morigen, Molina F, Skarstad K. Deletion of thesite does not affect once-per-cell-cycle timing but induces rifampin-resistant replication., 2005, 187(12): 3913–3920.
[14] Morigen, Løbner-Olesen A, Skarstad K. Titration of theDnaA protein to excesssites causes destabilization of replication forks, delayed replication initiation and delayed cell division., 2003, 50(1): 349–362.
[15] Oshima T, Aiba H, Masuda Y, Kanaya S, Sugiura M, Wanner BL, Mori H, Mizuno T. Transcriptome analysis of all two-component regulatory system mutants ofK-12., 2002, 46(1): 281–291.
[16] Ansaldi M, Simon G, Lepelletier M, Méjean V. The TorR High-Affinity Binding Site Plays a Key Role in BothAutoregulation andoperon Expression in., 2000, 182(4): 961–966.
[17] Selbach BP, Pradhan PK, Dos Santos PC. Protected sulfur transfer reactions by theSuf system., 2013, 52(23): 4089–4096.
(责任编委: 何群)
The effects of TorR protein on initiation of DNA replication in
Yuan Yao, Jiaxin Qiao, Jing Li, Hui Li, Morigen
The two-component systems, which could sense and respond to environmental changes, widely exist in bacteria as a signal transduction pathway. The bacterial CckA/CtrA, ArcA/ArcB and PhoP/PhoQ two-component systems are associated with initiation of DNA replication and cell division, however, the effects of the TorS/TorR system on cell cycle and DNA replication remains unknown. The TorS/TorR system incan sense changes in trimethylamine oxide (TMAO) concentration around the cells. However, it is unknown if it also affects initiation of DNA replication. We detected DNA replication patterns in Δand Δmutant strains by flow cytometry. We found that the average number of replication origins (s) per cell and doubling time in Δmutants were the same while the average number ofs in Δmutantswas increased compared with that in wild-type cells. These results indicated that absence of TorR led to an earlier initiation of DNA replication than that in wild-type cells. Strangely, neither overexpression of TorR nor co-expression of TorR and TorS could restore Δmutant phenotype to the wild type. However, overexpression of SufD in both wild type and Δmutants promoted initiation of DNA replication, while mutation of SufD delayed it in Δmutants. Thus, TorR may affect initiation of DNA replication indirectly through regulating gene expression of.
TorS/TorR two-component system; the response regulator TorR; DNA replication;
2014-09-01;
2014-09-30
国家自然科学基金项目(编号:31360208),内蒙古自然科学基金重大开放项目(编号:20102009)和内蒙古大学人才引进科研启动基金资助项目(编号:SPH-IMU, Z20090107)资助
姚远,在读博士研究生,专业方向:生物化学与分子生物学。E-mail:zhjyq129@163.com
莫日根,博士,教授,博士生导师,研究方向:生物化学与分子生物学,DNA复制与细胞周期。E-mail: morigenm@life.imu.edu.cn
10.16288/j.yczz.14-291
2015-1-5 10:50:43
http://www.cnki.net/kcms/detail/11.1913.R.20150105.1050.001.html
猜你喜欢
小猕猴智力画刊(2021年6期)2021-08-05
文化学刊(2019年12期)2019-12-26
中国设备工程(2019年24期)2019-01-16
现代园艺(2017年21期)2018-01-03
黑龙江史志(2016年9期)2017-03-16
中国民族医药杂志(2016年5期)2016-05-09
作文大王·低年级(2016年3期)2016-03-11
中国康复理论与实践(2015年10期)2015-12-24
医学研究杂志(2015年5期)2015-06-10
江苏教育·中学教学版(2015年3期)2015-05-13
