
胡杨种子萌发对温光条件和盐旱胁迫的响应特征
2015-07-04 09:22王瑞清李志军
西北植物学报 2015年8期
张 肖,王瑞清,李志军*
(1 新疆生产建设兵团 塔里木盆地生物资源保护利用重点实验室,新疆阿拉尔843300;2 塔里木大学 植物科学学院,新疆阿拉尔843300)
大多数植物生长都是从种子开始的,种子萌发阶段是植物体最脆弱的阶段[1],也是植物体重建至关重要的阶段[2,3],同时对植物繁殖及种群维持、扩展和恢复有着重要的意义[4]。种子萌发状况受到自身因素和环境条件的影响[1,5,6]。外界环境因子能够调节种子萌发的季节以及萌发的分布,从而有利于出苗和幼苗的建立[7]。干旱荒漠区降水稀少,蒸发强烈并伴随有土壤盐碱化,荒漠植物常常以其特殊的种子萌发机制来确保在合适的时间与地点完成种子萌发与幼苗生长发育,从而保证在极端严酷的环境条件下生存[8]。不同物种以及同一物种不同生态型的种子萌发对温度有不同的需求[9-14],变温促进种子萌发[6,15],影响着植物种子休眠的打破、萌发速率和最终萌发率,温度是影响种子萌发的关键环境因素之一[2,5]。对种子萌发温度的敏感性分析能够更好地预测温度对植物生活周期重要阶段的潜在影响[16]。在干旱半干旱荒漠地区,光照不是制约种子萌发的主要因素[5],大多数荒漠植物种子无论在光 照条件下还是在黑暗条件下都能萌发[9,10,13-15,17],只有一小部分植物种子萌发需要严格的光照[18-21]。光照可以通过影像种子的萌发影响植物的分布[7,12]。盐对植物的影响通过渗透胁迫降低植物从外界吸收水分的能力和离子积累的毒害作用而影响植物的生长[22]。水是荒漠植物种子萌发依赖的最重 要环境因子[5,6,8,19],降水或洪水漫灌可以增大土壤中水分的含量,降低土壤中的盐分[23],从而有利于种子的萌发。
胡杨(P.euphratica)隶属于杨柳科(Salicaceae)胡杨亚属,为起源古老的孑遗植物,是唯一能在干旱荒漠区自然形成大面积森林的乔木树种,为中国首批确定的388种珍稀濒危植物中的渐危种[24]。全世界60%的胡杨分布在中国,中国89.1%的胡杨主要分布在新疆塔里木河流域[25],塔里木河流域天然胡杨林对抵御风沙、遏制沙漠扩展、保护和改善自然环境、维持绿洲农业生产安全起到了至关重要的作用。胡杨种子在塔里木盆地散种时间为7月底至9月中旬,种子散布期间平均气温24.3 ℃、干燥地表平均温度约46.2 ℃、潮湿地表温度约35.4 ℃。胡杨实生苗主要出现在河漫滩,其他自然生境实生苗较少[26,27]。这是由于胡杨种子萌发对水分条件十分敏感,随着沙子水分含量的降低,种子萌发率迅速降低。当沙子含水量为27%时,种子的萌发率为86%;沙子含水量降为20%时,只有32%的种子能够萌发;在含水量降为13%后,种子已经不能从沙子中吸收水分,萌发率为0[28,29]。由洪水流量决定的河漫滩含水量,以及短暂洪水期后强烈地表蒸发引起的盐分在土壤表层积累[30],都成为胡杨种子萌发及幼苗早期成活的重要生态影响因子,盐分(土水比)越高种子萌发率越低[28],低浓度的盐就对种子的萌发有促进作用[4,31,32]。寿命短暂的胡杨种子[28]在自然条件下是如何响应温度、光照及盐旱胁迫而实现在河漫滩上进行种子繁殖的?本项研究在人工控制条件下探讨温度、光周期、干旱胁迫和盐胁迫对胡杨种子萌发的影响,旨在阐明种子萌发与温度、光照、干旱胁迫和盐胁迫的关系,揭示胡杨种子萌发阶段占据生境资源的生态适应策略。
1 材料和方法
1.1 材 料
本试验材料胡杨种子来源于新疆生产建设兵团农一师9团人工胡杨林(E 81°10.303′,N 40°33.434′,海拔904m)。采样地地处塔里木河上游,年均降水量45.5mm,蒸发量2 045mm;7月底至9月中旬平均气温在16.4~28.4 ℃,平均地温为21.7~36.1 ℃。于2013年9月采集胡杨成熟蒴果放在室内通风处自然干燥,待果皮自然开裂后将其置于纱网上轻轻揉搓,使冠毛和种子分离。将收集后的种子置于棕色瓶里密封,贮藏在4 ℃条件下备用。
1.2 试验设计
1.2.1 温光两因素实验 温度梯度处理为:10/15℃、15/20 ℃、20/25 ℃、25/30 ℃、30/35 ℃和35/40℃,光照处理为:连续光照、连续黑暗和12h黑暗/12h光照,共组成18个处理组合。
1.2.2 干旱胁迫试验 参考Michel[33]配制渗透势梯度为-0.1、-0.2、-0.4、-0.6、-0.8、-1.0、-1.2、-1.4和-1.6 MPa的处理溶液,以蒸馏水为对照。在环境条件为12h 光照/12h 黑暗、25℃/30 ℃(黑暗/光照)的培养箱中进行种子萌发期干旱胁迫试验。
1.2.3 盐胁迫试验 试验设置0.05、0.10、0.20、0.30、0.40、0.60和0.80mol/L的NaCl处理溶液,以蒸馏水为对照,在环境条件为12h光照/12h黑暗、25 ℃/30 ℃(黑暗/光照)的培养箱中进行种子萌发期盐胁迫试验。
1.3 种子萌发试验和观察指标
种子萌发试验是在RTOT 系列人工智能气候箱中进行。实验前用75%的酒精对培养皿进行擦拭消毒。将种子置于直径为90 mm 垫有2层湿润滤纸的培养皿中培养,每处理种子100 粒,重复3次。每12h统计1次萌发种子数,种子萌发以胚根出现为标志。温光试验持续7d,干旱胁迫和盐胁迫持续4d。根据统计结果计算不同处理条件下种子的最终萌发率(GF)、萌发速率(GI)和平均萌发时间(MGT),公式如下:
最终萌发率(GF)=∑n/N
萌发速率(GI)=∑100n/ND
平均萌发时间(MGT)=∑(Dn)/∑n
式中,N是供试种子数,D是从萌发开始的时间,n是在时间D 的种子萌发数。
1.4 数据分析
利用DPS7.55软件对数据进行方差分析,采用LSD 法进行多重比较。
2 结果与分析
2.1 温度和光照对胡杨种子萌发的影响
表1显示在3种光照条件下,胡杨种子最终萌发率在15 ℃/20 ℃~35 ℃/40 ℃范围内均达到90%以上,均在25 ℃/30 ℃~30 ℃/35 ℃达到最大值,且显著高于10 ℃/15 ℃处理;在20 ℃/25 ℃~30 ℃/35 ℃范围,连续光照(L)、连续黑暗(D)和12 h光照/12h黑暗(L/D)3种光照处理之间胡杨种子最终萌发率无显著差异。
3种光照条件下,胡杨种子萌发速率在10 ℃/15 ℃~30 ℃/35 ℃范围随温度升高而升高,在25℃/30 ℃~35 ℃/40 ℃范围种子萌发速率均达到最高,且显著高于10 ℃/15 ℃~20 ℃/25 ℃范围的种子萌发速率(表1);同时,在20 ℃/25 ℃~30 ℃/35℃范围,连续光照(L)、连续黑暗(D)和12h光照/12h黑暗(L/D)处理间种子萌发速率无显著差异。
3种光照条件下,胡杨种子平均萌发时间在10℃/15 ℃~30 ℃/35 ℃温度范围内随温度升高而降低,其中25 ℃/30 ℃~35 ℃/40 ℃范围种子平均萌发时间明显低于10 ℃/15 ℃~20 ℃/25 ℃范围的平均萌发时间,在该温度范围3种光照条件下各温度之间平均萌发时间均无显著差异。在同一变温条件下,种子平均萌发时间均以连续黑暗(D)条件下最低,而另外两种光照条件下较高且相近,除20℃/25 ℃、30 ℃/35 ℃温度处理外,光照处理间种子平均萌发时间均存在显著性差异(表1)。同时,在25℃/30 ℃~35 ℃/40 ℃温度范围内胡杨种子起始萌发率较高(>50%),且在48h以内基本都能完成种子萌发,而在15 ℃/20 ℃~35 ℃/40 ℃温度范围,胡杨种子均可以在72h内完成萌发(图1)。
综合以上结果表明,胡杨种子在3种光照处理下所有的试验温度范围均可以萌发,最适萌发温度范围在25 ℃/30℃~30 ℃/35 ℃之间,在此温度范围内3种光照条件对种子萌发的影响无显著差异,而且在此温度范围种子萌发表现出快速且集中的特点。
2.2 干旱胁迫对胡杨种子萌发的影响
根据上述光温实验结果,在12h光照/12h黑暗、25℃/30℃条件下进行PEG6000模拟干旱胁迫试验。结果发现,胡杨种子最终萌发率在溶液渗透势为-0.1~-1.0 MPa范围内时均大于80%,且各处理及对照间均无显著差异;但当PEG6000溶液渗透势由-1.00 MPa降到-1.20 MPa时,最终萌发率则迅速大幅下降;随着溶液渗透势继续下降,最终萌发率则降至10%以下,直至为0(图2,A)。同时,渗透势0~-0.2 MPa PEG6000溶液对胡杨种子萌发进程无明显影响,表现在初始12h种子累积萌发率就达到65%以上,在24h内基本完成萌发,其萌发速率也与对照无显著差异;但溶液渗透势在-0.2~-1.4 MPa范围时,初始萌发率随着渗透势的降低逐渐降低,其萌发速率也显著逐渐降低,-1.2和-1.4 MPa处理最终累计萌发率已降至20%以下,萌发速率均在1%/h以下;-1.6MPa处理始终无种子萌发(图2,B、C)。以上结果表明,胡杨种子萌发可以忍受一定的渗透胁迫,但渗透胁迫超过一定范围会对种子萌发产生抑制;当渗透势低于-0.4 MPa时胡杨种子萌发进程和萌发速率受到显著影响,当溶液渗透势低于-1.2 MPa时种子萌发受到显著抑制。

表1 温度和光周期对胡杨种子萌发的影响Table1 The influence of three light conditions and temperature treatments on the seed germination of P.euphratica

图1 同一光照条件下温度对胡杨种子萌发进程的影响Fig.1 The influence of temperature on the final germination percentage of P.euphraticaseeds under the same light condition
2.3 盐胁迫对胡杨种子萌发的影响
根据光温实验结果,在12h光照/12h黑暗、25℃/30 ℃条件下进行NaCl胁迫试验。结果发现,在0~0.80mol/L 浓度范围内,胡杨种子最终萌发率随着NaCl溶液浓度的增高逐渐降低,但其在0~0.2mol/L 内均达到80%以上,而在0.40 mol/L NaCl浓度下开始大幅度降低至20%以下,在0.80 mol/L NaCl溶液浓度则下降到2.27%(图3,A)。同时,胡杨种子初始萌发率随着NaCl溶液浓度的增高逐渐降低、萌发高峰期逐渐向后推移,萌发速率降低。其中,当NaCl溶液浓度在0~0.1 mol/L时,种子萌发高峰期出现在12h内,累积萌发率均达到60%以上,萌发速率在6%/h以上;当NaCl浓度在0.30~0.80mol/L下时,种子萌发高峰期出现在12~24h内,萌发速率降至3%/h以下(图3,B、C)。以上结果表明胡杨种子萌发对盐胁迫具有一定的耐性,但在NaCl溶液浓度高于0.20mol/L 时就受到显著影响。

图2 渗透胁迫对胡杨种子最终萌发率(A)、萌发速率(B)和萌发进程(C)的影响Fig.2 The influence of osmotic stress on the germination percentage(A),germination rate(B)and germination process(C)of P.euphratica
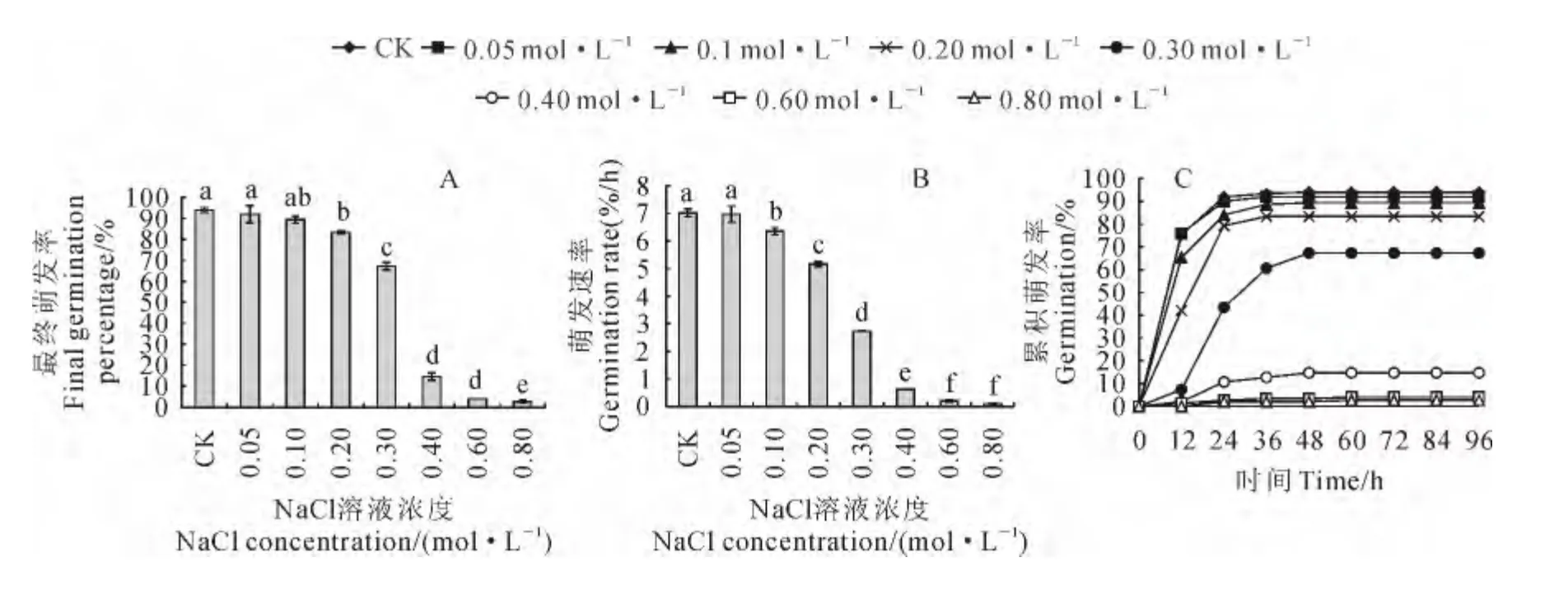
图3 盐胁迫对胡杨种子最终萌发率(A)、萌发速率(B)和萌发进程(C)的影响Fig.3 The influence of salt stress on the germination percentage(A),germination rate(B)and germination process(C)of P.euphratica
3 讨 论
3.1 胡杨种子萌发对温度、光照的响应特征
在荒漠或半荒漠地区,植物种子的萌发对光照有不同的要求,大部分种子萌发对光照不敏感,但部分荒漠植物种子的萌发需要严格光照,如Artemisiamonosperma,Capparisdeciduas,白沙蒿(Artemisiasphaerocephala)[18-20]。有些植物种子要在暗中萌发,如沙芥属的沙芥和斧翅沙芥种子在适宜的萌发温度和水分条件下,光成为抑制种子萌发的重要因子,并随光强和光照时间的增加对种子萌发有显著的抑制作用[13],而碟果虫实(Corispermumpatelliforme)种子只有在连续黑暗的条件下才能萌发[14]。大多数荒漠植物种子的萌发无论在光下还是在暗中都萌发得很好,如梭梭种子无论在光下和暗中均能萌发,萌发率无显著性差异[17]。本试验研究发现,胡杨种子在连续光照、连续黑暗和12h光照/12h黑暗下都能萌发,且具有较高的萌发率(在10 ℃/15 ℃条件下最低萌发率达到77.67%);不同温度条件下各光照处理间最终萌发率、萌发速率和平均萌发时间相差都很小,表现出对光照不敏感。胡杨种子极小、寿命短,当种子被风力传播到水分含量瞬息万变的河漫滩上,只要捕捉到适宜的含水量则不受光照条件的限制而迅速萌发,无疑增加了种子繁殖成功的机率。已有研究和试验观察都表明,胡杨种子在水分适宜的条件下4~6h即开始萌发,6~12h子叶开始展平接受光照而很快进入幼苗的早期生长阶段[26,31,32]。
不同的植物种子需要不同的萌发温度,其最适萌发温度也不同,这和其生活习性是相适应的。种子萌发对温度的要求确保了大部分种子在合适的季节萌发,可以增大幼苗存活的机会[7]。温度强烈影响种子的萌发率、萌发速率和萌发进程,适宜温度可促进种子萌发和幼苗生长[2,5]。本研究表明,胡杨种子在10 ℃/15 ℃~35 ℃/40 ℃下都能萌发,最终萌发率可以达到77%以上,萌发温度范围比较宽泛,这与李利等研究发现胡杨种子在恒温10℃~40℃时萌发率均超过50%结果一致[32]。另外,本研究还发现在25 ℃/30 ℃~35 ℃/40 ℃温度范围内胡杨种子萌发比较快速而且集中。在干旱荒漠区自然条件下,昼夜存在较大的温度变化,较宽泛的种子萌发温度范围是胡杨长期适应环境温度变化规律的结果,能够满足随风传播到适宜土壤水分环境的种子随时可以萌发,从而充分利用空间中有限的水分。另外,胡杨种子具有较高的最适萌发温度范围,同样是对胡杨果实成熟期7~9月地表温度较高的适应。反过来,较高的温度促使种子快速而集中萌发,种子萌发对温度的要求可以确保种子在昼夜不同温度下随时萌发,因而增大了幼苗的存活机会。这是胡杨种子利用温、光的宽泛条件在尽可能短的时间内占有和利用有效资源的另外一条生态适应策略。在适宜的温度下,种子萌发过程迅速,可以在1~2d完成萌发。分析认为,胡杨种子能够在较宽的温度范围内萌发,并迅速完成萌发过程,且不受光照条件限制,这些特点弥补了其种子寿命短的缺陷,充分利用短暂适宜的萌发条件,对胡杨天然更新有着重要的现实意义,是种子萌发阶段快速占据生境资源的生态适应策略之一。
3.2 胡杨种子萌发对干旱胁迫的响应特征
水势同温度一样是控制种子萌发的基本环境因子,对种子最终萌发率和萌发速率都有影响[2]。外界环境渗透势的大小决定着种子细胞从外界吸收水分的能力,外界渗透势越小种子吸收水分的难度越大,对低渗环境的适应也是种子萌发策略的一种适应。本研究发现,胡杨种子在渗透势为-1.2 MPa下溶液中可以萌发,与Li等[34]研究结果一致。胡杨种子萌发率在0~-1.0 MPa渗透势胁迫下大于90%,相互之间无显著差异,其耐渗透胁迫能力与梭梭(Haloxylonammodendron)相比偏低,但比碱蓬(Suaedaglauca)[35]强,而与刚毛柽柳(Tamarix hispida)[36]相当。本实验中,当外界渗透势在0~-0.2 MPa间,胡杨种子萌发进程基本不受影响,且12h萌发率均达到65%以上,24h 基本完成种子萌发。这些都说明胡杨种子可以忍受一定程度的渗透胁迫,从而利用有限的水资源而快速萌发。同时,胡杨虽然能在低渗溶液中萌发,但子叶展开和胚根伸长都受到影响,只有在-0.2 MPa以上时种子才能正常的生长,这也是只能在河漫滩地才能看到胡杨实生小苗[26,27],而在其他地方则很少能见到的原因。胡杨种子寿命短暂[28],干旱荒漠地区降水稀少,蒸腾强烈,胡杨散种时间与河流洪水期一致[26],胡杨种子高渗透势下快速萌发对有限水资源的利用具有重要意义。
3.3 胡杨种子萌发对NaCl胁迫的响应特征
盐对植物的影响主要表现在渗透效应和离子效应两个方面。前者影响植物对水分的吸收,后者在植物中积累到一定水平后造成毒害作用。盐对种子萌发的影响主要表现在降低种子萌发率,延长种子萌发进程,或者使种子失去活力[9,11,17]。杨帆等[9]对角果碱蓬(Suaedacorniculata)二型性种子(棕色和黑色)研究发现,盐胁迫对两类型种子的生活力均无影响,但盐胁迫显著降低两类型种子的初始萌发率、恢复萌发率和最终萌发率。而黄振英等[17]研究发现梭梭种子在浓度低于0.2mol/L 的NaCl溶液下萌发受到影响不大,但从0.8mol/L 起其萌发率随着浓度增高而降低,且NaCl处理后的部分种子永久地失去萌发力。本研究发现,胡杨种子萌发在NaCl溶液浓度高于0.20mol/L 时受到显著影响,种子最终萌发率降低,这与刘建平等[31]和王海珍等[37]研究结果一致,同时我们还发现NaCl胁迫降低了种子萌发速率,推迟了种子的萌发高峰出现时间。刘建平等和王海珍等研究还发现随着盐浓度的增加胡杨子叶展叶率呈显著下降趋势,不利于子叶的同化作用,影响植株的建立。对种子萌发时的盐胁迫研究最能反映该植物的耐盐性,胡杨林在中国主要分布在海拔800~1 100m 之间的荒漠内陆河流沿岸冲积平原上[25],而自然条件下胡杨实生苗主要分布在河漫滩地,一方面水是胡杨种子萌发的主要限制因子之一,另一方面也有可能是对盐的一种规避。河水漫灌后降低了土壤中的盐分[23],从而有利于胡杨种子快速萌发。这也从侧面说明了与河流洪水期一致的胡杨散种时间选择是对干旱荒漠环境的一种适应。
[1] GUTTERMAN Y.Strategies of seed dispersal and germination in plants inhabiting deserts[J].TheBotanicalReview,1994,60(4):373.
[3] DODD G L,DONOVAN L A.Water potential and ionic effects on germination and seedling growth of two cold desert shrubs[J].American JournalofBotany,1999,86(8):1 146-1 153.
[5] REN J(任 珺),YU F K(余方可),TAO L(陶 玲).Research advances on the germination of desert plants under stress[J].Bulletinof BotanicalResearch(植物研究),2011,31(1):121-128(in Chinese).
[6] CHANG W(苌 伟),WU J G(吴建国),LIU Y H(刘艳红).Research advance in seed germination of desert woody plants[J].Chinese JournalofAppliedEcology(应用生态学报),2007,18(2):436-444(in Chinese).
[7] ZHANG J N(张佳宁),LIU K(刘 坤).Mechanisms for plants detecting the optimum time and place to germinate[J].ActaPrataculturaeSinica(草业学报),2014,23(1):328-338(in Chinese).
[8] ZHANG Y(张 勇),XUE L G(薛林贵),GAO T P(高天鹏),etal.Research advance on seed germination of desert plants[J].Journalof DesertResearch(中国沙漠),2005,25(1):106-112(in Chinese).
[9] YANG F(杨 帆),CAO D CH(曹德昌),YANG X J(杨学军),etal.Adaptive strategies of dimorphic seeds of the desert halophyteSuaedacorniculatain saline habitat[J].ChineseJournalofPlantEcology(植物生态学报),2012,36(8):781-790(in Chinese).
[10] MA H G(马海鸽),JIANG Q(蒋 齐),WANG ZH J(王占军),etal.Effects of light and temperature on seed germination ofGlycyrrhiza uralensisunder different pretreatment[J].ResearchofSoilandWaterConservation(水土保持研究),2014,21(5):225-229,235(in Chinese).
[11] GAO R R(高瑞如),ZHAO R H(赵瑞华),ZHANG SH F(张双风),etal.Effects of salt and temperature onHalocnemumstrobilaceumseed germination[J].ActaBotanicaBoreali-OccidentaliaSinica(西北植物学报),2007,27(11):2 281-2 285(in Chinese).
[12] SOCOLOWSKI F,VIEIRA D C M,TAKAKI M.Interaction of temperature and light on seed germination inTecomastansL.Juss.ex Kunth(Bignoniaceae)[J].BrazilianArchivesofBiologyandTechnology,2008,51(4):723-730.
[13] SONG ZH W(宋兆伟),HAO L ZH(郝丽珍),HUANG ZH Y(黄振英),etal.Effects of light and temperature on the germination ofPugioniumcornutum(L.)Gaertn.andPugioniumdolabratumMaxim.seeds[J].ActaEcologicaSinica(生态学报),2010,30(10):2 562-2 568(in Chinese).
[14] LIU Y J(刘有军),JI Y F(纪永福),MA Q L(马全林),etal.Effect of light and temperature on the germination of three annual plants[J].ChineseJournalofEco-Agriculture(中国生态农业学报),2010,18(4):810-814(in Chinese).
[15] LIU L CH(刘龙昌),SHEN F(沈 芳),MAIMAITIJIANG(买买提江),etal.Seed germination inNanophytonerinaceumand the spatial distribution of its seedlings[J].ActaBotanicaBoreali-OccidentaliaSinica(西北植物学报),2007,27(3):607-611(in Chinese).
[16] WALCK J L,HIDAYATI S N,DIXON K W,etal.Climate change and plant regeneration from seed[J].GlobalChangeBiology,2011,17(6):2 145-2 161.
[17] HUANG ZH Y(黄振英),ZHANG X SH(张新时),GUTTERMAN Y,etal.Influence of light,temperature and salinity on the seed germination ofHaloxylonammodendron[J].ActaPhytoecologicaSinica(植物生理学报),2001,27(3):275-280(in Chinese).
[18] HUANG Z Y,GUTTERMAN Y.Influences of environmental factors on achene germination ofArtemisiasphaerocephala,a dominant semishrub occurring in the sandy desert areas of Northwest China[J].SouthAfricanJournalofBotany,1999,65:187-196.
[19] HUANG Z Y,GUTTERMAN Y.Comparison of germination strategies ofArtemisiaordosicawith its two congeners from deserts of China and Israel[J].ActaBotanicaSinica,2000,42(1):71-80.
[20] QAISER M,QADIR S A.A contribution to the autecology ofCapparisdecidua(Forssk.)EdgewⅠ.Seed germination and the effect of topographic conditions on the growth,abundance and sociability[J].PakistanJournalofBotany,1971,3(1):37-60.
[21] SOCOLOWSKI F,VIEIRA D C M,SIMAO E,etal.Influence of light and temperature on seed germination ofCereuspernambucensisLemaire(Cactaceae)[J].BiotaNeotropica,2010,10(2):53-56.
[22] MUNNS R.Comparative physiology of salt and water stress[J].Plant,CellandEnvironment,2002,25:239-250
[23] FU J Y(傅荩仪),XU H L(徐海量),ZHAO X F(赵新风),etal.Differences in the impacts of flooding frequency and duration on riparian vegetation and soil in the Lower Tarim River[J].ActaPrataculturaeSinica(草业学报),2013,22(6):11-20(in Chinese).
[24] SU P X(苏培玺),ZHANG L X(张立新),DU M W(杜 明武),etal.Photosynthetic character and water use efficicency of different leaf shapes ofPopuluseuphraticaand their response to CO2enrichment[J].ActaPhyoecologicaSinica(植物生态学报),2003,27(1):34-40(in Chinese).
[25] WANG SH J(王世绩).The status,conservation and recovery of global resources ofPopuluseuphradica[J].WorldForestryResearch(世界林业研究),1996,6:38-45(in Chinese).
[26] HUA P(华 鹏).Studies on Seed germination and seeding early growth ofPopuluseuphraticaon The Flood Plain[J].Environmental ProtectionofXinjiang(新疆环境保护),2003,25(4):14-17(in Chinese).
[27] ZHAO ZH Y(赵振勇),ZHANG K(张 科),LU L(卢 磊),etal.Seedling recruitment in desert riparian forest following river flooding in themiddle reaches of the Tarim River[J].ActaEcologicaSinica(生态学报),2011,31(12):3 322-3 329(in Chinese).
[28] ZHANG Y B(张玉波),LI J W(李景文),ZHANG H(张 昊),etal.Analysis on the factors cause the failure ofPopuluseuphraticasextual regeneration in nature[J].ScienceTechnologyandEngineering(科学技术与工程),2005,5(8):467-472(in Chinese).
[29] 李俊清,卢 琦,褚建民,等.额济纳绿洲胡杨林研究[M].北京:科学出版社,2009.
[30] FOUZIA B A,JANIS S M.Germination and seedling survival of Argem(Arganiaspinosa)under experimental saline conditions[J].JournalofAridEnvirornmnents,2001,49:533-540.
[31] LIU J P(刘建平),LI ZH J(李志军),HE L R(何良荣),etal.Salt-tolerance ofPopuluseuphraticaandP.pruinosaSeeds during Germination[J].ScientiaSilvaeSinicae(林业科学),2004,40(2):165-169(in Chinese).
[32] LI L(李 利),ZHANG X M(张希明),HE X Y(何兴元).Effects of Salinity and water potential on seed germination and radicle growth of riparianPopuluseuphratica[J].AridZoneResearch(干旱区研究),2005,22(4):520-525(in Chinese).
[33] MICHEL B E,KAUFMANN M R.The osmotic potential of polyethylene glycol 6000[J].PlantPhysiology,1973,51(5):914-916.
[34] LI L,ZHANG X M,MICHAEL R,etal.Responses of germination and radicle growth of twoPopulusspecies to water potential and salinity[J].ForestryStudiesinChina,2006,8(1):10-15.
[35] YANG J N(杨景宁),WANG Y R(王彦荣).Effects of drought stress simulated by PEG on seed germination of four desert plant species[J].ActaPrataculturaeSinica(草业学报),2012,21(6):23-29(in Chinese).
[36] SUN H Y(孙红叶),LI L(李 利),LIU G J(刘国军),etal.Effects of temperature,salt and water stress on the seed gemination ofTamarixhispida[J].AridLandGeography(干旱区地理),2007,30(3):414-420(in Chinese).
[37] WANG H ZH(王海珍),HAN L(韩 路),JIA W S(贾文锁).Response of seeds germination ofPopuluseuphraticaandPopuluspruinosato slat stress[J].JournalofDesertResearch(中国沙漠),2013,33(3):743-750(in Chinese).
猜你喜欢
中国机械工程(2022年8期)2022-05-09
金桥(2021年11期)2021-11-20
儿童时代·幸福宝宝(2021年1期)2021-03-29
北广人物(2020年47期)2020-12-09
中学生数理化·高一版(2020年6期)2020-07-25
小资CHIC!ELEGANCE(2019年40期)2019-12-10
大众文艺(2019年17期)2019-09-09
创新作文(1-2年级)(2018年6期)2018-01-22
中学生数理化·八年级物理人教版(2017年3期)2017-11-09
信息安全与通信保密(2016年3期)2016-08-23
