
秦岭南坡松栎林群落演替过程中种间联结性和相关性研究
2015-07-04 09:22张岗岗王得祥张明霞刘文桢郭小龙
西北植物学报 2015年8期
张岗岗,王得祥*,张明霞,刘文桢,郭小龙
(1 西北农林科技大学 林学院,陕西杨陵712100;2 甘肃小陇山森林生态系统国家定位观测研究站,甘肃天水741020;3 青海省社会科学院,西宁810000;4 小陇山林业科学研究所,甘肃天水741020)
植物群落是由在时间和空间上彼此相关联的共存物种组成,不同时空各物种之间的相互关系决定着群落的结构特征和动态[1]。研究种间关系不仅可以反映群落内物种的数量特征,更重要的是能有效反映群落中各物种对环境因子的适应程度及特定环境因子作用下种间相互依存和制约关系,对于正确认识群落演替过程中种间替代和共存关系演变机制具有重要意义[2-5]。种间联结是把成对物种的存在与否作为两个物种出现的相似性的定性特征,种间相关则是一种定量关系,二者通常是由群落生境的差异影响了物种的分布和生长而引起的,反映了不同物种在时间与空间上的相互关系[6-8]。基于种间关系的生态种组是从强联结的种对中提炼出来,反映了种对在生态过程中的异质性和物种生态需求的趋同性,也是研究种间关系的基础[9],对其进行划分有助于客观地了解区域植物群落的结构,并有助于探讨不同物种之间、物种与环境间关系,为物种多样性的保护、自然植被的恢复与重建提供理论依据[4-5,10]。国内外学者对不同群落种间关系和生态种 组划分进行了大量研究[4-5,10-14],但这些研究主要针对某一特定群落或群落某一演替阶段优势种间关系进行定性分析的,缺乏对不同演替阶段群落种间关系的定性和定量双重探讨,在生态种对层面划分生态种组的研究也鲜见报道。
在海拔800~2 300m 之间的秦岭山地森林生态系统中,油松林(Pinustabulaeformisforest)、锐齿栎林(Quercusalienavar.acuteserrataforest)及其混交林 (P.tabulaeformis-Q.alienavar.acuteserratamixed forest)是该区3 种典型的森林群落类型[15,16]。国内学者关于秦岭地区油松林、松栎混交林和锐齿栎林这3个群落是否处于同一演替序列中的不同阶段还存在争议,但部分学者认为松类树种是先锋种,松栎混交林是松类林受到干扰后整个群落处在被栎类植物逐渐取代的初级演替阶段,而栎类的竞争能力比较强,其取代油松的趋势十分明显[17,18],而对这3个群落更新特征的研究也表明油松林群落终将为锐齿栎林群落所取代[19-23]。最近,部分学者单独对这3种群落进行更新特征、生态位特征、种间联结等方面研究[9,11,19-24],张子良等[15]、张洪武等[16]则分别探讨了这3种群落乔灌草层物种组成与环境因子、物种多样性之间的关系,但针对秦岭林区油松林→松栎混交林→锐齿栎林这一演替序列过程中种间关系动态变化研究尚属空白。秦岭松栎林演替过程中不同演替阶段群落优势种的总体联结性如何变化?主要种群种间联结性的显著程度及大小如何?群落演替过程中种间相关性如何?这些问题和规律的深入研究和探讨,有助于阐明不同演替阶段物种之间替代和共存关系,并为推演秦岭林区松栎次生林演替趋势奠定基础。鉴于此,本研究采用空间替代时间的方法,选取秦岭南坡典型的油松林、松栎混交林、锐齿栎林为研究对象,阐述了不同演替阶段群落种间联结性和相关性动态变化规律,以期揭示优势种群种间关系和群落演替的相互作用,为秦岭山地松栎林演替过程中健康稳定的群落关系构建提供科学依据。
1 研究区概况
研究区域为秦岭南坡中段主要包括秦岭南坡至汉水以北的汉中市各(区)县以及与之相邻的凤县部分地区,即E105°30′~108°11′,N32°50′~33°53′。该区地形地貌极为复杂,全年雨量充沛,气候温暖湿润,差异明显,海拔800 m 以下为北亚热带,800~2 400m为山地暖温带,2 400 m 以上为高山寒温带。本研究具体地点是以火地塘、旬阳坝等为主的5个松栎林典型分布区域。
秦岭火地塘林区位于秦岭南坡中山地带中部(33°25′~33°29′N,108°25′~108°30′E),海拔800~2 500m,年均气温8~10 ℃,年均降雨量900~1 200mm,年日照时数1 327.5h,属中国北亚热带和暖温带的过渡地带。以山地棕壤、暗棕壤和山地草甸土为主,土层厚30~50cm,该区属温带针阔混交林和寒带针叶林,从低海拔到高海拔依次为:松栎林亚带,松桦林亚带,云、冷杉针阔叶混交林亚带。主要树种有:锐齿栎(Quercusalienavar.acuteserrata)、红 桦(Betulaalbo-sinensis)、油 松(Pinus tabulaeformis)、华山松(Pinusarmandii)等。旬阳坝林区位于秦岭中段南麓的月河梁和平河梁之间(32°29′~33°13′N,103°58′~109°48′E),平均海拔1 300m,年均气温为10 ℃,年均降水量1 133mm,年日照时数1 638.3h,该区属北亚热带温暖湿润气候区。土壤为矿砾质壤黏土,呈微酸性,该区属暖温带落叶阔叶林和针阔混交林向北亚热带常绿落叶混交林的过渡带,林木主要有松、杉和落叶乔木等。
2 研究方法
2.1 野外调查
采用空间代替时间的方法,在对油松林→松栎混交林→锐齿栎林这一演替序列分布较集中的火地塘、旬阳坝等林区全面踏查的基础上,选择样地时尽可能缩小各阶段间的空间距离,并尽量保持立地因子一致、人为干扰较小的自然演替群落,每个群落类型(阶段)采用典型取样法设置重复样地15个,样地面积为20m×20m,在每个样地四角和中心分别设置5个2m×2m 灌木样方和1m×1m 草本样方。对样地内所有乔木进行每木检尺、坐标定位,记录其物种名称、胸径、树高、冠幅和生长状况等;灌草样方记录种类、株数、盖度、分布和生长状况等;详细记录海拔、坡度、坡向、坡位等立地因子。45个调查样地基本情况,即样地编号、经纬度、海拔、坡度、坡向、坡位、郁闭度、林龄、土壤等立地条件均已在参考文献[15,16]中详细列出,本文不再赘述。
2.2 数据统计与分析
分别计算秦岭南坡油松林、松栎混交林、锐齿栎林各15个样地乔木层物种重要值的平均值,列表统计重要值排序前20的种群(表1)在各样方中出现与否的1、0二元数据矩阵,采用方差比率、χ2检验、2×2联列表、Jaccard指数、Spearman秩相关系数等分析方法,分别对松栎林不同演替阶段群落中主要种群间总体相关性、种对间的联结性、种对间联结强度和种间相关性进行计算、统计和分析。具体方法参见文献[3,6-8,25]。
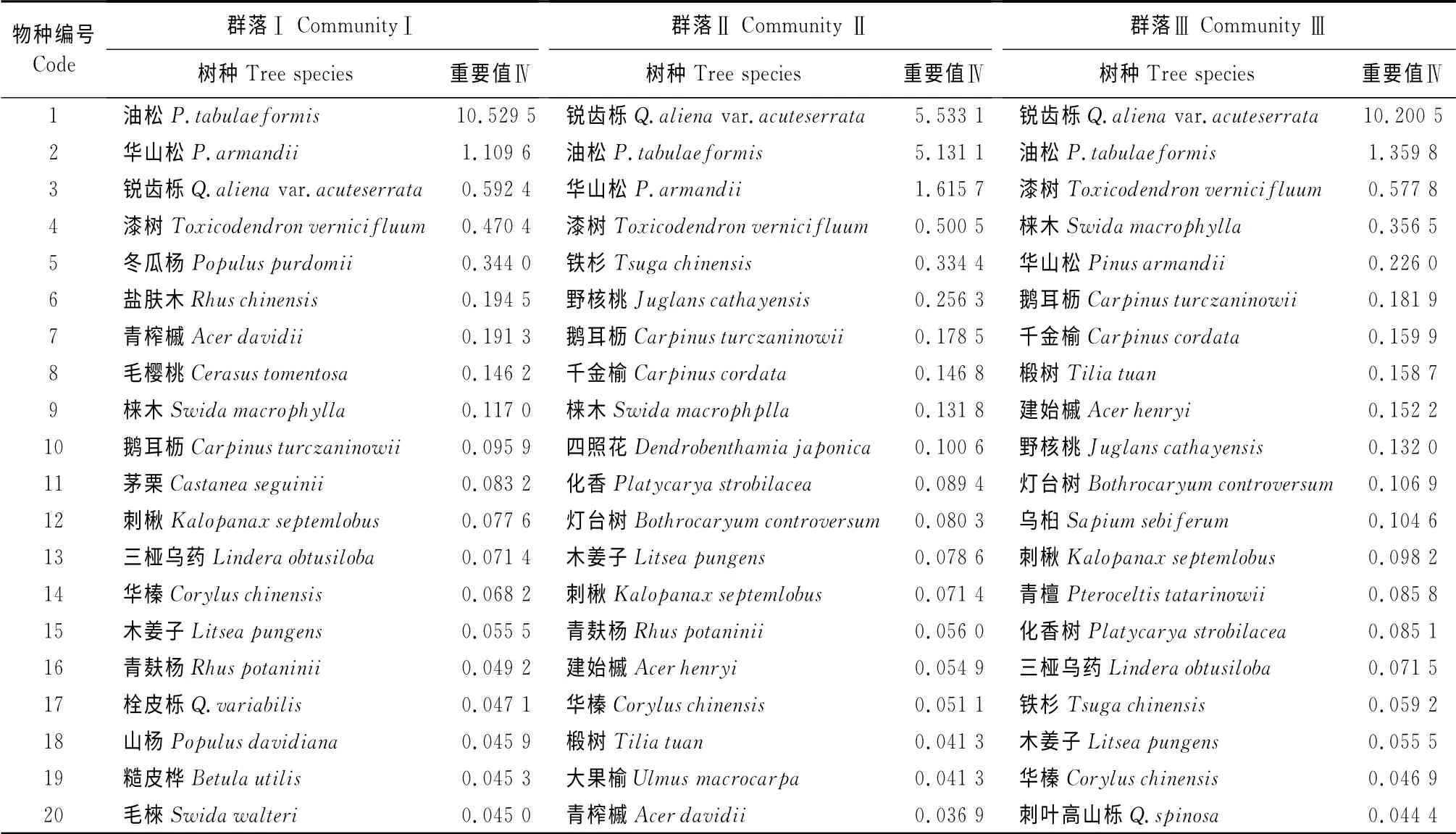
表1 秦岭南坡不同演替阶段3种森林群落物种组成及重要值Table1 Importance values and species components of three forest communities at different stages of succession on the south-facing slopes in Qinling Mountains
3 结果与分析
3.1 群落不同演替阶段优势种总体关联性分析
群落中优势树种间的总体联结性反映了群落内各树种间关联性的总体趋势。不同演替阶段优势种间的总体联结性分析结果(表2)表明,油松林、松栎混交林和锐齿栎林优势种间的方差比率分别为1.81、1.97、2.45,均大于1,表明在独立零假设条件下,总体联结性表现为净的正联结;显著性χ2检验统计量W 分别为27.14、29.61、36.73,落在区间(7.261,27.996)外,说明群落总体的正联结程度均显著,且随演替进程而加强,因此,不同演替阶段的群落均表现为显著正联结,种间关系紧密联系,各群落具有相对的稳定性,也侧面反映了主要种群所在的林区大环境与各群落生境需求的一致性,使其更易组成群落并保持其稳定性。
3.2 群落不同演替阶段主要种对联结性分析
种间联结的χ2检验能比较准确地反映出种间联结的显著程度,JI值则能体现出χ2检验不显著种对联结性的大小,其值越接近1,表明该种对的正联结越紧密,其值越接近0,则表明负联结越强。

表2 秦岭南坡松栎林群落不同演替阶段群落优势种总体关联性Table2 The overall interspecific connectivity of dominant species in P.tabulaeformis-Q.alienavar.acuteserrata mixed forest communities at different stages of succession on the south-facing slopes in Qinling Mountain
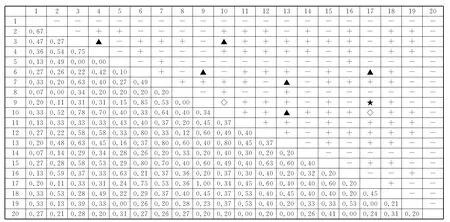
图1 秦岭南坡油松林群落主要种对间χ2 检验和JI半矩阵图Fig.1 The semi-matrix diagram aboutχ2 test and JI values of dominant species in P.tabulaeformis forest communities on the south-facing slopes in Qinling Mountain
在油松林群落中(图1),主要种群χ2检验中85对为正关联,占总种对数的44.7%,其中极显著正关联的有1对,显著正关联的有6对;有103对为负关联,占总对数的54.2%,均不显著;有2对为无关联,群落中正关联的种对数明显小于负关联,显著或极显著的种对仅占4.7%,为锐齿栎-漆树、青榨槭-三桠乌药、鹅耳枥-三桠乌药、梾木-栓皮栎等,这些种对生态学特性相近,生境需求相似,种间关系协同进化,趋于稳定,其余大多数种对则处于不显著水平,说明油松群落种间联结性较松散,这与总体联结性分析结果一致。群落中JI>0.75的有9对,0.75≥JI>0.5的有28对,约占总种对数的14.7%;0.5≥JI>0.25的有97对,约占总对数的51.1%;0.25≥JI≥0的有56对,约占总种对数的29.5%。进一步说明个别种对间对资源和空间存在一定的竞争,但不激烈,多数种对种间关联程度不紧密,这与χ2检验结果相似。因此,油松林群落整体上处于较为稳定的阶段。
松栎混交林群落中(图2),主要种群χ2检验中有81对为正关联,占总对数的42.6%,其中显著正关联3对,分别为华山松-漆树、漆树-鹅耳枥、野核桃-鹅耳枥;有106对为负关联,占总对数的55.6%,其中显著负关联的1对,为漆树-灯台树;其余3对为无关联,正负关联比为0.76,绝大多数种对处于不显著水平,说明松栎混交林群落中多数种对间存在微弱的相互竞争或依赖关系。群落中JI>0.75的仅有8对,占总种对数的4.2%;0.75≥JI>0.5的有21对,约占总种对数的11.1%;0.5≥JI>0.25的有113对,占总种对数的59.5%;0.25≥JI≥0的有48对,约占总种对数的25.3%,说明群落中多数种对联结程度较弱,群落结构和状态相对稳定。值得注意的是,χ2检验结果表明锐齿栎、油松、华山松、漆树之间正联结关系并不显著,但JI值远大于0.75,说明这些树种之间为显著或极显著正关联,因此,为避免χ2检验结果产生偏差,引入JI值测定种间关联程度是必要的。
锐齿栎林群落中(图3),主要种群χ2检验中有98对为正关联,占总对数的51.6%,其中显著正关联2对,为鹅耳枥-千金榆、建始槭-刺楸;有88对为负关联,占总对数的55.6%,均不显著;有4对为无关联,正负关系比为1.11,检验显著率仅为1.05%,大部分种对间正或负关联性不显著,对资源和空间的竞争较为缓和,种间关系趋于稳定共存,群落总体处于比较稳定的状态。群落中JI>0.75的种对仅有4 对,漆树-华山松、梾木-鹅耳枥也呈显著正关联;0.75≥JI>0.5的有31对,占总对数的16.3%;0.5≥JI>0.25的有118对,占总种对数的62.1%;0.25≥JI≥0的有37对,占总种对数的19.5%。说明大多数种对间的关联程度不高,物种分布具有一定的独立性,群落稳定性较高。
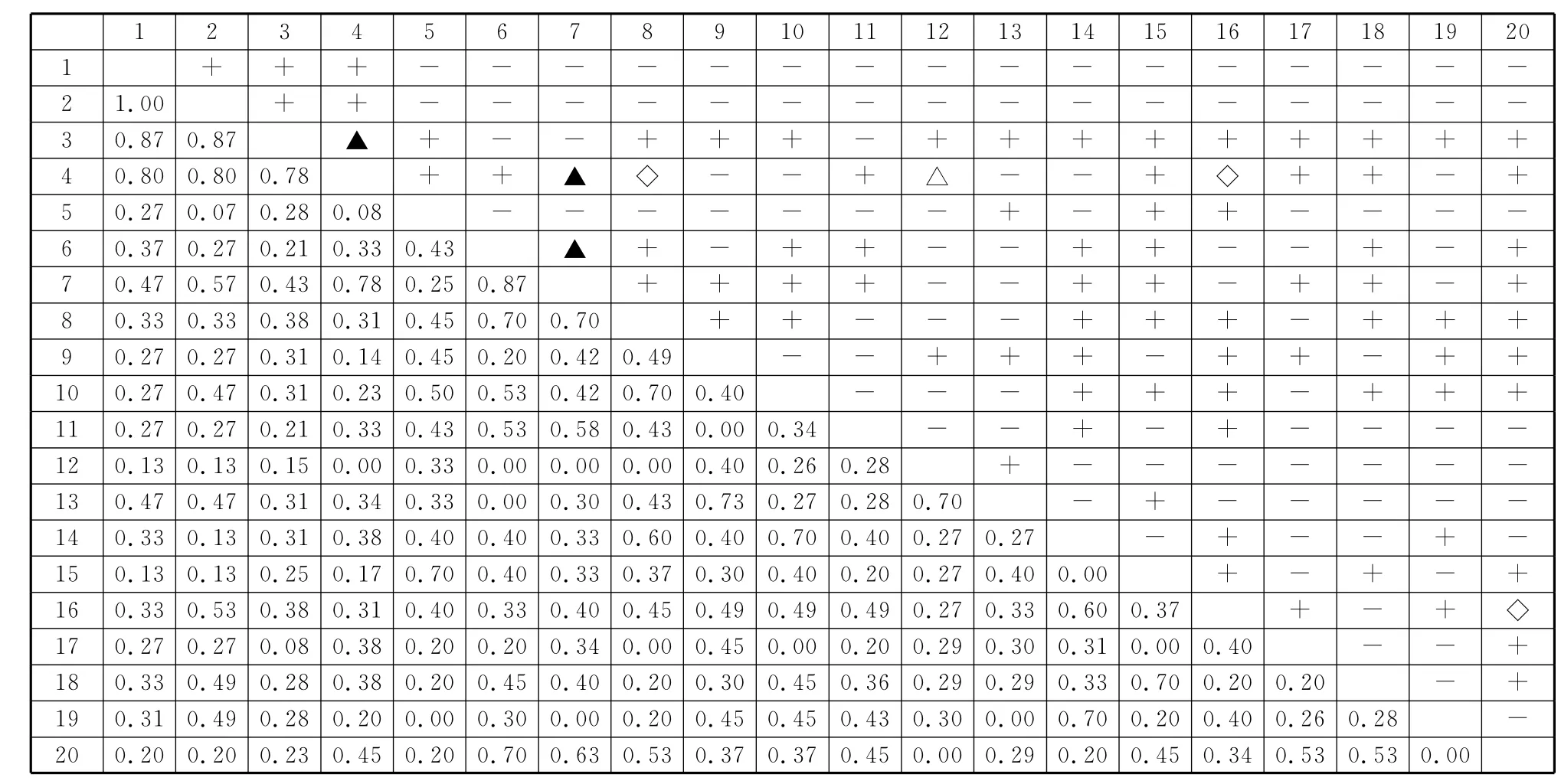
图2 秦岭南坡松栎混交林群落主要种对间χ2 检验和JI半矩阵图Fig.2 The semi-matrix diagram aboutχ2 test and JI values of dominant species in P.tabulaeformis-Q.aliena var.acuteserrata mixed forest communities on the south-facing slopes in Qinling Mountain
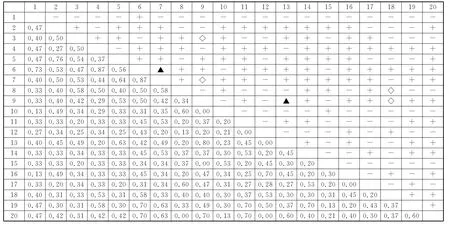
图3 秦岭南坡锐齿栎林群落主要种对间χ2 检验和JI半矩阵图Fig.3 The semi-matrix diagram aboutχ2 test and JI values of dominant species in Q.alienavar.acuteserrata forest communities on the south-facing slopes in Qinling Mountain
综上所述,油松林→松栎混交林→锐齿栎林这一演替序列过程中,前期和中期群落物种间正联结比例接近,分别为44.7%和42.6%,后期增加到51.6%;负联结比例从54.2%略增大到55.6%后减小至46.3%;均有少量无关联种对出现,说明随着演替的进行,群落中主要种群对生态要求逐渐趋于相似;前期和中期群落中达到显著或极显著水平的正关联种对占有一定比例,均多于后期,负关联中达到显著水平及以上的种对极少;JI≥0.5和0.25≥JI≥0的比例逐渐减少,0.5≥JI>0.25逐渐增多,表明群落演替过程中种间关联程度趋于弱度联结,因此,群落在长期的协同进化过程中种间关系逐渐趋于稳定共存。其次,χ2检验中显著正关联种对JI值都比较高,显著负关联种对的JI值较小,即JI在反映种对之间关联性和关联程度方面与χ2检验结果相一致,与郭垚鑫等[13]研究结果基本一致。
3.3 不同演替阶段物种之间的联结性分析
根据相关研究[26]及演替特性[27],将各演替阶段重要值排名前20的乔木划分为4个树种组,其中油松、华山松、山杨、冬瓜杨、糙皮桦为先锋树种,青榨槭、青麸杨、梾木、鹅耳枥、刺楸、榛子、木姜子、三桠乌药为前期过渡种,灯台树、化香树、榛子、梾木、鹅耳枥、千金榆、椴树、建始槭、刺楸、榛子、木姜子、三桠乌药为后期过渡种,锐齿栎、漆树、野核桃、铁杉为顶级树种。由演替过程中各树种组组内及组间种对联结性统计结果(表3)可知,先锋树种之间正联结和负联结比例分别为33.3%和66.7%,均不显著,华山松与油松之间竞争较为激烈,JI值达0.67,总体处于不稳定阶段;先锋种与前期过渡种之间正关联种对占38.9%、负关联种对占61.1%,正负关联比增大,种间关系趋于紧密;随着演替过程中的激烈竞争,前期过渡种间、后期过渡种间及其前期过渡种和后期过渡种之间表现为较为有利的正联结,分别占各自总种对数的59.2%、50.0%和53.4%,虽然正负联结比均大于1,但不明显,处于群落动态演替过程中相对稳定的阶段;由于顶级树种的侵入和竞争,后期过渡种与顶级种之间正关联比例明显下降,负关联比例高达63.4%,不同树种之间对资源和空间实行再分配;顶级种间正联结比例仅为22.2%,负关联比例达77.8%,说明这些顶级树种之间竞争较为激烈,尚未完全分化协调、演替至稳定阶段。整个演替过程中不同物种之间不断竞争资源和空间,物种组成及其种间关系处于动态变化过程中,群落结构也将不断完善并趋于稳定。
3.4 群落不同演替阶段主要种对相关性分析
Spearman秩相关系数是检验2个物种线性关系的重要指标,其处理的是定量数据,反映2个物种同时出现的可能程度[3]。不同演替阶段群落优势种间Spearman秩相关性分析结果中(图4~图6),油松林群落正相关种对为88对,占总对数的46.3%,其中极显著正相关8对,显著正相关4对;负相关种对有100对,占总对数的52.6%,其中,极显著负相关1对,显著负相关3对;无关种对有2对;正负相关比为0.88,检验显著率为6.32%。松栎混交林群落正相关种对为89对,占总对数的46.8%,其中极显著正相关4 对,显著正相关14 对;负相关种对100对,占总对数的52.6%,其中,极显著负相关4对,显著负相关3对,无关种对1对;正负相关比为0.89,检验显著率为13.16%。锐齿栎林群落正相关种对为99对,占总对数的52.1%,其中极显著正相关9对,显著正相关5对;负相关种89对,占总对数的46.8%,其中,显著负相关的有1对;无关种对有2对;正负相关比为1.11,检验显著率为7.89%。由此可知,与锐齿栎林群落相比,油松林和松栎混交林群落种间正负相关比接近,且均明显小于锐齿栎林,群落中多数物种间的关系不紧密,这与χ2检验和JI半矩阵分析结果基本一致,这符合前人研究结果[3,28];显著率则先增大后减小,但均大于χ2检验
结果,主要是因为利用χ2检验来判断种间关联性会损失一定的信息量[28],而Spearman秩相关系数检验灵敏度较高,可以弥补χ2检验的不足,这与之前研究结果相似[28,29]。

表3 秦岭南坡松栎林群落演替过程中先锋种、过渡种以及顶级种间的种间联结Table3 The interspecific associations of pioneer species,transitionary species and climax species in P.tabulaeformis-Q.alienavar.acuteserrata mixed forest communities at different stages of succession on the south-facing slopes in Qinling Mountain

图4 秦岭南坡油松林群落优势种间Spearman秩相关系数半矩阵图Fig.4 The semi-matrix diagram about Spearman’s rank correlation coefficient of dominant species in P.tabulaeformis forest communities on the south-facing slopes in Qinling Mountain
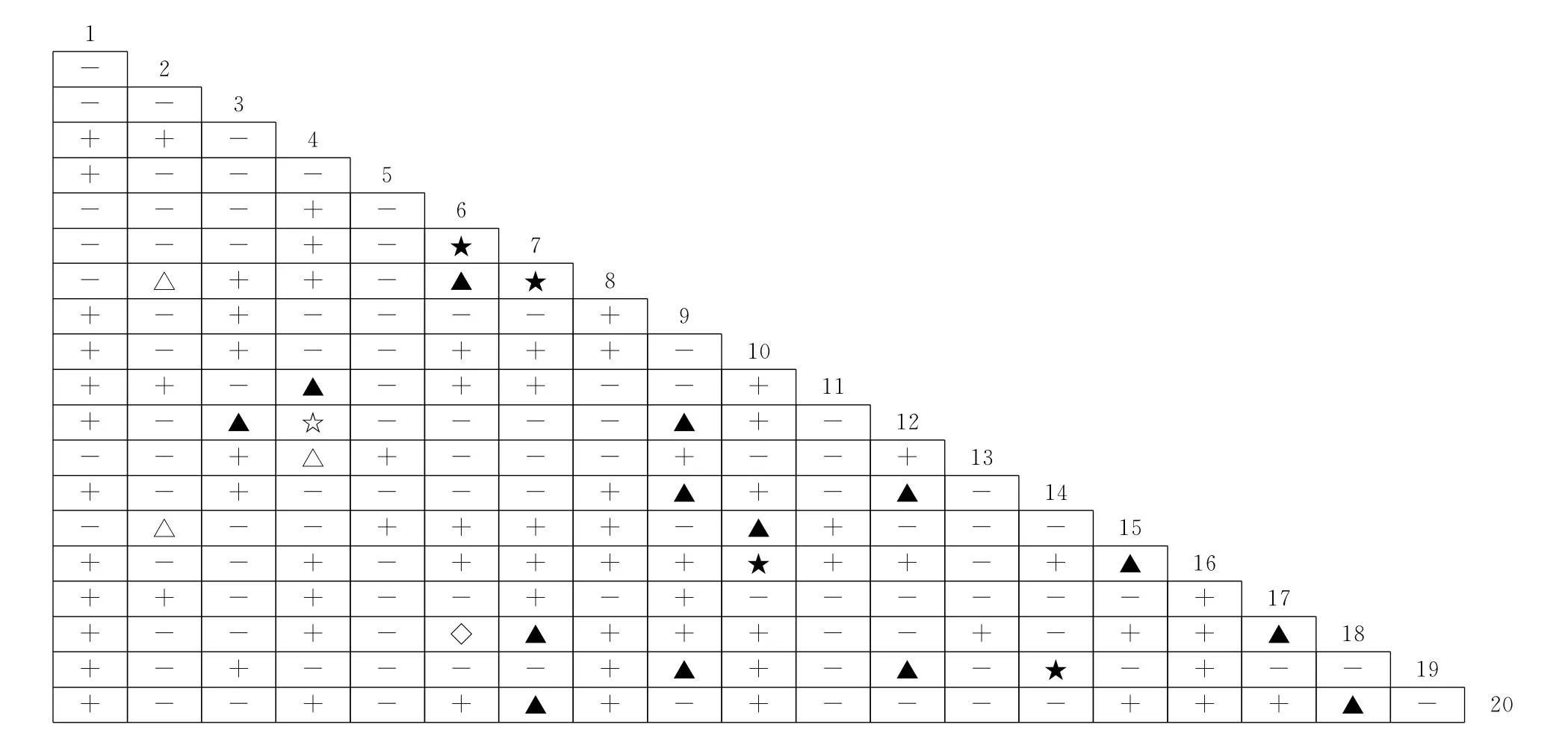
图5 秦岭南坡松栎混交林群落优势种间Spearman秩相关系数半矩阵图Fig.5 The semi-matrix diagram about Spearman’s rank correlation coefficient of dominant species in P.tabulaeformis-Q.alienavar.acuteserrata mixed forest communities on the south-facing slopes in Qinling Mountain
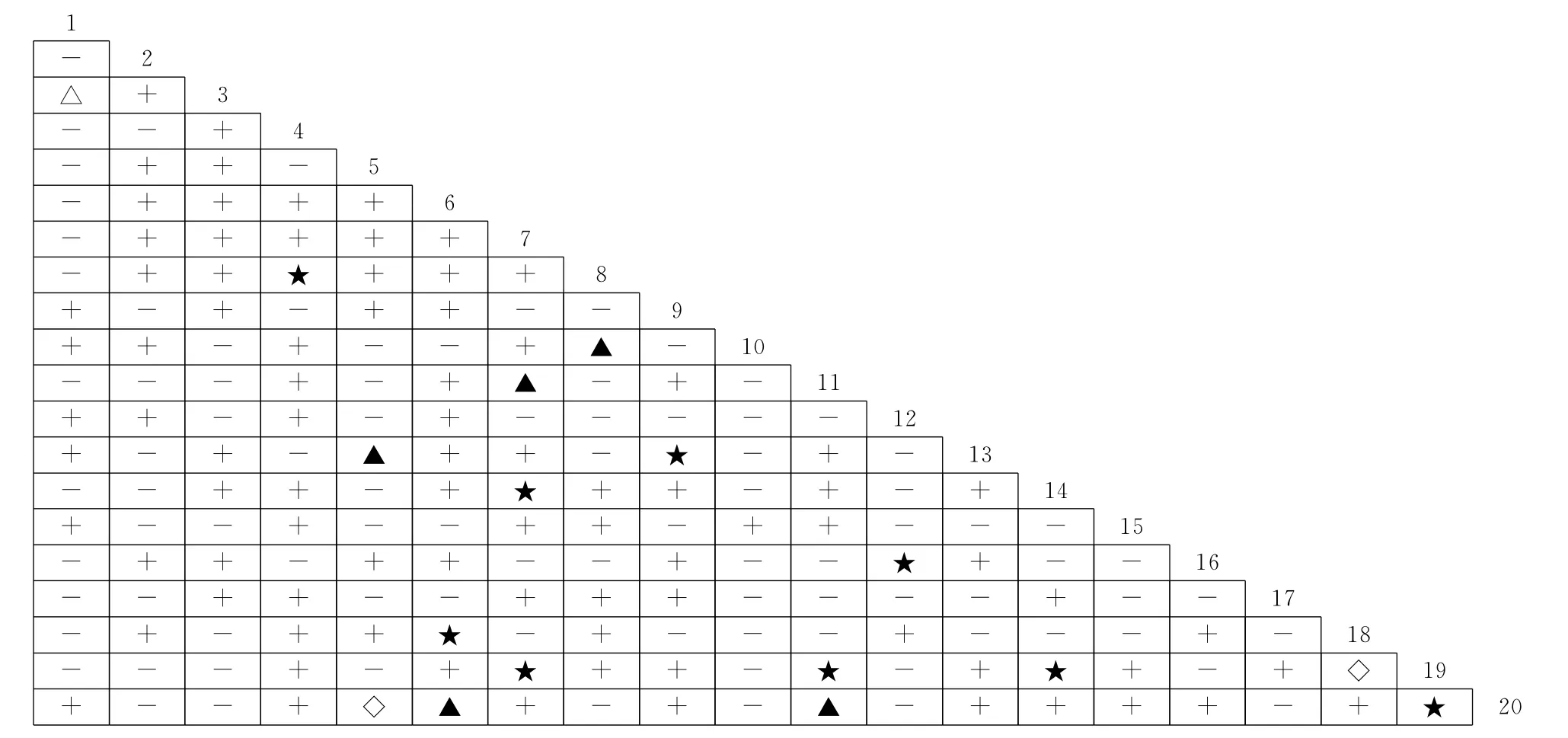
图6 秦岭南坡锐齿栎林群落优势种间Spearman秩相关系数半矩阵图Fig.6 The semi-matrix diagram about Spearman’s rank correlation coefficient of dominant species in Q.alienavar.acuteserrataforest communities on the south-facing slopes in Qinling Mountain
3.5 群落不同演替阶段生态种组划分
为了更真实可靠地反映群落种间关联以及群落演替方向,依据不同演替阶段群落种间关联和相关性计算结果,以显著负相关性为界限,同一生态种组内的物种有尽可能大的正相关性和关联性,物种之间具有相似的生态要求和生活习性[2,29],将不同演替阶段群落生态种组进行划分。油松林群落第1生态种组为华山松、冬瓜杨、山杨、油松,其中油松和华山松为主要建群种,存在一定的竞争,但生态习性相似,华山松与冬瓜杨、山杨则呈显著正关联和相关性;第2生态种组为刺楸、盐肤木、梾木、鹅耳枥、三桠乌药、青榨槭、木姜子,处于群落的中下层,为主要伴生种,种间关联性和相关性较显著;第3生态种组为锐齿栎、栓皮栎、漆树、糙皮桦、榛子、微毛樱桃,多为阳生树种,分布广,联结不明显,但相关性较显著;第4生态种组为青麸杨、毛梾、毛栗,处于劣势,彼此间相互独立。松栎混交林群落第1生态种组为锐齿栎、油松、华山松、铁杉,为群落主要优势树种,两两之间多为正关联且JI值极大;第2生态种组为野核桃、千金榆、鹅耳枥、椴树、青榨槭、华榛,数量较多,为主要伴生树种,正相关性较显著,多为不显著正关联;第3生态种组为化香、漆树,二者之间呈不显著正关联和正相关;第4生态种组为灯台树、梾木、大果榆、刺楸,为林下次要伴生树种,相关性较显著,多与其它树种呈不显著负关联;第5生态种组为四照花、建始槭、青麸杨、木姜子,数量较少,随机分布,独立性较强。锐齿栎林群落第1生态种组为锐齿栎、油松、漆树、华山松,为群落主要优势种,多为正关联,JI值极大;第2生态种组为梾木、椴树、野核桃,为主要伴生种,竞争上层资源和空间,多为显著正相关和不显著正关联且JI值较大;第3生态种组为华榛、千金榆、灯台树、青檀、刺叶栎、木姜子、鹅耳枥、乌桕、三桠乌药、刺楸、建始槭,为中下层优势种群,彼此间多为极显著正相关和不显著正关联且JI值较小;第4生态种组为铁杉、化香树,数量较少,独立性较强。
4 结论与讨论
4.1 种间联结与群落稳定性
秦岭南坡松栎林群落不同演替阶段群落总体关联性均为显著正关联,各自具有相对的稳定性,且总体关联性随着演替进程而加强,稳定性也逐渐增强,这与长白山云冷杉针阔混交林演替过程中总体联结性随演替发展的变化规律一致[30]。前人对秦岭林区松栎林种间关系表明,秦岭林区油松天然林大多数种群种间联结很弱,具有相对独立分布的特征[9],华山松天然次生林优势种群间联结较松散,物种之间存在一定的独立性,趋于形成一个较为稳定的群落[11-28],锐齿栎林绝大多数种对间的独立性相对较强,群落处于较为稳定的次生林演替阶段[12-13],这些不同演替阶段特定群落类型总体均表现出一定的松散性,但独立程度较低,本研究结果与之较为一致。这种种间联结的松散性可能主要决定于物种的生物学和生态学特性[4,7-8],同时受群落演替的时间和空间[11]及其生态位分化的影响,此外,外界干扰和病虫害等也会影响群落结构的稳定性。
4.2 种间联结与群落演替的关系
一般来说,随着植被群落演替的进展,群落结构及种类组成将逐渐趋于完善和稳定,种间关系也将同步趋于正关联,以求得多物种间的稳定共存[25,31]。秦岭南坡松栎林演替过程中主要植物种间联结性随演替进展所表现出的特点也遵循这一规律,与油松林和松林混交林群落相比,锐齿栎林群落物种间正联结比例较高,这与物种对群落的适应性和群落对物种的选择和淘汰密切相关[11,30],但检验显著率和JI>0.75或0.25≥JI≥0的比例较低,趋于相对独立分布,稳定性明显高于演替前期和中期。Spearman秩相关性研究结果进一步证实油松林和松栎混交林群落中多数物种间的关系不紧密,锐齿栎群落种间关系则达到某种协调,生态位彼此分化表现出明显的正相关特性,表明群落在长期的协同进化过程中种间关系趋于稳定共存。前人对中国森林演替模式的研究表明,次生演替一般遵循针叶林到 针阔混交林再到阔叶林的规律[7,32,33],就种间关系来说,本研究结果支持这一演替规律,但仅从种间联结和相关性来反映秦岭南坡松栎林群落演替动态是不够全面的,今后还需结合群落结构、更新特征等进一步分析考证。
对油松林→松栎混交林→锐齿栎林这一演替序列整个过程中的先锋树种、前期过渡种、后期过渡种和顶级种种间联结性研究发现,种对之间的正负关联比先增大后减小,近似呈单峰分布,即油松、华山松等先锋树种在群落中均占优势,且生态习性和生境需求相似,表现出对资源和空间的激烈竞争,正负联结种对分别占总种对数的33.3%和66.7%,尚未形成稳定的群落关系,必然导致林木间出现自疏和他疏,形成林隙,林下充足的光照、水热、养分和生存空间有利于青榨槭、鹅耳枥、木姜子、千金榆、椴树等阔叶树种的侵入和定居,重新形成激烈的种间竞争格局,经过长期的生态位分化,种间关系趋于松散,因此,这些过渡种间(包括前期过渡种间、后期过渡种间及其相互之间)多表现为有利的正联结,正负联结比均大于1,但不明显,处于群落动态演替过程中相对稳定的阶段;而适应性强且生长迅速的顶级树种锐齿栎、漆树、野核桃等阔叶树种则与后期过渡种间的负关联比例达63.4%,经过不断的竞争和淘汰逐渐占据上层空间,相同的空间分布和生长资源竞争使得这些树种之间竞争较为激烈,多形成显著的负联结,正负关联比仅为0.29,而鹅耳枥、千金榆等主要伴生树种则在林分中下层占有一定的优势,与上层顶级种间多形成较为广泛的不显著正关联,正是这些不同层次和不同生活型物种间对资源和空间需求的差异,使得物种组成及其种间关系处于动态变化过程中,群落结构也将不断完善并趋于稳定。马映栋等[13]对锐齿栎林种群空间分布格局和关联性研究也证实了这一点,林冠上层锐齿栎分布稳定,主要为随机分布,林间层多均匀分布,林下层聚集分布明显,膀胱果(Staphyleaholocarpa)与锐齿栎在幼树期因相互竞争而大部分尺度上互为负相关,其生长发育到一定阶段相互促生且小部分尺度上相互独立;树种锐齿栎与青榨槭、膀胱果与青榨械在整体上保持无关联,逐渐发展为小范围内互为相关性。群落中种间关系是复杂的,是多种因子耦合的结果[34],仅仅依靠同一时空尺度下的调查资料和种对联结统计结果分析,尚不能完全揭示松栎林演替过程中的种间关系,更需要对演替过程中多物种的联结关系进行长期定位研究,同时考虑环境因子和人为因素的影响,才能更好揭示群落种间相互关系和演替趋势。
4.3 种间联结与物种多样性关系
物种多样性是反映植物群落组成结构和稳定性的重要指标,种间关系是群落形成、演化和稳定性的重要数量特征[35],二者之间存在一定的联系。对油松林→松栎混交林→锐齿栎林这一演替序列整个过程中的先锋树种、前期过渡种、后期过渡种和顶级种种间联结性研究发现,种对之间的正负关联比先增大后减小,近似呈单峰分布,也即先锋种定居后,青榨槭、鹅耳枥等阔叶树种的侵入促使种间竞争更加激烈,而过渡种间关系趋于松散,顶级树种锐齿栎、漆树、野核桃等竞争也较为激烈。而秦岭南坡松栎林群落演替过程中虽然群落间物种丰富度无显著差异,但群落总体物种多样性随演替进展总体上呈现明显的单峰型变化,在松栎林阶段物种多样性达到最大,与中期物种多样性假说一致[16]。这与沙棘人工林群落物种多样性及种间关联变化特征较为一致,沙棘人工林群落各层及总体物种多样性指数基本呈“S”形波动变化的趋势,种间关联正负关联比呈抛物线形变化,说明群落发育到13a前后呈现相对稳定的态势[35]。这可能是因为群落经过阔叶树种入侵过程的激烈竞争与淘汰,种间关系则达到某种协调,生态位彼此分化,群落组成和结构达到了稳定共存的阶段。
4.4 种间关系与种群生态位关系
种间联结与其生态位重叠之间是紧密联系的。油松林中锐齿栎-油松、漆树-华山松、锐齿栎-漆树、盐肤木-栓皮栎、锐齿栎-鹅耳枥χ2检验值较大,呈显著正关联或一定的正关联,其生态位重叠值相对较大[9,24],分 别 为0.589、0.561、0.231、0.189、0.176,而鹅耳枥-梾木、栓皮栎-鹅耳枥呈显著负关联,其生态位重叠值[9]仅为0.033、0。锐齿栎林中锐齿栎-鹅耳枥、华山松-油松、华山松-鹅耳枥、漆树-油松、漆树-三桠乌药、油松-鹅耳枥、三桠乌药-鹅耳枥等呈一定程度的正关联,其生态位重叠值[36]分别为0.881、0.892、0.956、1.00、0.971、1.00、0.836;锐齿栎-椴树、锐齿栎-华山松、锐齿栎-漆树、鹅耳枥-漆树、椴树-华山松等呈一定的负关联,其生态位重叠值[37]仅为0.441、0.502、0.301、0.467、0.317,由此可见,优势种群间的正联结性越强,其生态位重叠值就越大;反之,种间的负联结性越强,其生态位重叠值就越小,这与前人的研究结果一致[38,39]。
4.5 种间联结与CCA 排序关系
通过对比种间联结性及其种对在物种与环境因子CCA 排序图[15]中的分布可以发现,锐齿栎林中的锐齿栎与鹅耳枥在CCA 排序图中的距离较近,而与椴树、华山松、漆树相距较远;鹅耳枥与锐齿栎相距较近,而与漆树相距较远;漆树与油松相距较近,而与锐齿栎、鹅耳枥相距较远。松栎混交林中的漆树与华山松、鹅耳枥相距较近,而与油松、铁杉相距较远。由此可见,这些种对在物种与环境因子CCA 排序图中分布距离远近很大程度上取决于其正负关联性,也就是说,对于同一物种,若与其它物种间存在较为显著的正联结性,则正联结性越显著,物种在CCA 排序图上的距离越近;若与其它物种间存在较为显著的负联结性,则负联结性越显著,物种在CCA 排序图上的距离越远。这与郑超超等[39]对浙江江山公益林种间关系的研究结果较为一致。这可能是因为正关联种对能够充分利用周围资源空间,生态位重叠程度较高,导致其在CCA 排序图中集中分布,而负关联种对偏好于某些环境因子,整体上对环境资源的利用能力较差,因而分散分布于CCA 排序图中。此外,油松林关联种对间分布距离与其他2种群落类型有较大差异,这可能是因为油松林的分布与锐齿栎林和松林混交林在CCA 排序图中的分布相距较远,即其群落在结构组成和生境等方面差异较大[15]。CCA 排序同时也表明,坡度是影响乔木层物种分布的主要环境因子,其次是海拔、坡度等[15],正联结的物种生境特征比较一致,负联结的物种生境特征刚好相反。物种之间的关系也不完全受环境条件制约,还需进一步结合生态位理论、CCA 协变量矩阵偏典范对应分析等探讨。
4.6 种间联结对林分经营的指导
秦岭南坡油松林、锐齿栎林及其混交林稳定性程度直接关系到秦岭生物多样性维持、水碳平衡、气候调节、水土保持以及水源涵养等生态服务功能的发挥。阐释植物种群种间关系旨在揭示种对间竞争的结果或群落的现状,这有助于对地带性植被进行改造和恢复,有效缩短演替进程。油松林中锐齿栎-漆树、盐肤木-梾木,松栎林中的锐齿栎-油松、锐齿栎-漆树等呈显著正关联的种对应作为生态目标树或经营目标树予以保留;相反,油松林中的油松-三桠乌药、漆树-冬瓜杨,松栎混交林中的灯台树-漆树、锐齿栎-化香树等种对呈不显著负关联且JI值较大或显著负关联的种对应选择性的伐除;此外,还可依据种间关系的变动趋势确定抚育间伐时植株的去留[30]和采伐阶段,例如华山松-梾木种间联结性演替中期呈不显著正联结、前期和后期均为不显著负联结,应选择松栎混交林向锐齿栎林过渡阶段调整华山松和梾木的种间关系;生态种组的划分也为筛选群落内环境监测的指示种[28],满足林区营林和重建需要提供了保障,通过这些有效的经营措施促进林分向健康稳定的地带性顶级群落发展,缩短秦岭山地松栎林演替进程。
[1] ARMAS C,PUGNAIRE F I.Plant interactions govern population dynamics in semi-arid plant community[J].JournalofEcology,2005,93(5):978-989.
[2] LIU J H(刘珏宏),GAO H(高 慧),etal.Comparative analysis of interspecific association within theStipagrandis-S.kryloviicommunity in typical steppe of Inner Mongolia,China[J].ChineseJournalofPlantEcology(植物生态学报),2010,34(9):1 016-1 024(in Chinese).
[3] GUO Y X(郭垚鑫),HU Y N(胡有宁),KONG L T(孔令童),etal.Interspecific association and correlation of main plant species in subalpine meadow of Zibai Mountain[J].ActaEcologicaSinica(生态学杂志)2011,30(8):1 775-1 780(in Chinese).
[4] SU S J,LIU J F,HE Z S,etal.Ecological species groups and interspecific association of dominant tree species in Daiyun Mountain National Nature Reserve[J].JournalofMountainScience,2015,12(3):637-646.
[5] LI Y D,XU H,etal.Division of ecological species groups and functional groups based on interspecific association——a case study of the tree layer in the tropical lowland rainforest of Jianfenling in Hainan Island,China[J].FrontiersofForestryinChina,2008,3(4):407-715.
[6] GREIG-SMITH P.Quantitative Plant Ecology(3rd edition)[M].BlackwellScientificPublications,1983.
[7] PENG SH L(彭少麟),ZHOU H CH(周厚诚),GUO SH C(郭少聪),etal.Studies on the changes in interspecific association of zonal vegetation in Dinhushan[J].ActabotanicaSinica(植物学报),1999,41(11):1 239-1 244(in Chinese).
[8] WANG B S(王伯荪),PENG SH L(彭少麟).Study on the measuring techniques of interspecific association of lower-subtropical evergreenbroadleaved forests I.The exploration and the revision on the measuring formulas of interspecific association[J].ActaPhytoecologicaet GeobotanicaSinica(植物生态学与地植物学丛刊),1985,9(4):274-285(in Chinese).
[9] LIN Y Y(蔺雨阳),WANG D X(王得祥),LEI R D(雷瑞德),etal.Classification of ecological twin species and plant functional type for naturalPinustabulaeformisForests in middle part of Qinling Mountains[J].JournalofNortheastForestryUniversity(东北林业大学学报),2009,37(11):46-51(in Chinese).
[10] XU J SH(许金石),CHEN Y(陈 煜),WANG G X(王国勋),etal.Interspecies association of dominant woody plant communities in Qinling forest region,Shaanxi[J].ActaBot.Boreal.-Occident.Sin.(西北植物学报),2014,34(7):1 467-1 475(in Chinese).
[11] ZHANG M X(张明霞),WANG D X(王得祥),KANG B(康 冰),etal.Interspecific associations of dominant plant populations in secondary forest ofPinusarmandiiin Qinling Mountains[J].ScientiaSilvaeSinicae(林业科学),2015,51(1):12-21(in Chinese).
[12] YANG H Y(杨海裕),ZHANG S ZH(张宋智),etal.Interspecific associations among tree species in natural deciduous broadleaved forests in western Qinling,Northwest China[J].ChineseJournalofEcology(生态学杂志),2012,31(10):2 513-2 520(in Chinese).
[13] MA Y D(马映栋),LIU W ZH(刘文桢),etal.Change analysis of spatial pattern and correlation forQuercusalienavar.acuteserrataforest population in Xiaolongshan Mountains[J].ActaBot.Boreal.-Occident.Sin.(西北植物学报),2014,34(9):1 878-1 886(in Chinese).
[14] LIU Y Y,LI F R,JIN G Z.Spatial patterns and associations of four species in an old-growth temperate forest[J].JournalofPlantInteractions,2014,9(1):745-753.
[15] ZHANG Z L(张子良),YU F(于 飞),WANG D X(王得祥).Effects of environmental factors on the species composition of three typical secondary forests in Qinling Mountains[J].JournalofNorthwestA&FUniversity(Nat.Sci.Ed.)(西北农林科技大学学报·自然科学版),2014,42(8):69-78(in Chinese).
[16] ZHANG H W(张洪武),YU F(于 飞),WANG D X(王得祥),etal.Changes of species composition and diversity in the process of community succession of pine oak forest on the south-facing slopes in Qinling Mountains[J].ActaBot.Boreal.-Occident.Sin.(西北植物学报),2014,34(1):169-176(in Chinese).
[17] BARBÉRO M,LOISEL R,QUÉZEL P,etal.Pines of the Mediterranean Basin[M]//RICHARDSON D M.Ecology and Biogeography of Pinus,Cambridge University Press,Cambridge.1998:153-170.
[18] GRACIA M,RETANA J,ROIG P.Mid-term successional patterns after fire of mixed pine-oak forests in NE Spain[J].ActaOeclolgica,2002,(23):105-411.
[19] KANG B(康 冰),LIU SH R(刘世荣),WANG D X(王得祥),etal.Regeneration characteristics of woody plant seedlings in typical secondary forests in Qinling Mountains[J].ChineseJournalofAppliedEcology(应用生态学报),2011,22(12):3 123-3 130(in Chinese).
[20] YU F,WANG D X,SHI X X,etal.Seed dispersal by small rodents favors oak over pine regeneration in the pine-oak forests of the Qinling Mountain,China[J].ScandinavianJournalofForestResearch,2013,28(6):540-549.
[21] YU F,WANG D X,SHI X X,etal.Effects of environmental factors on tree seedling regeneration in a pine-oak mixed forest in the Qinling Mountains,China[J].JournalofMountainScience,2013,10(5):845-853.
[22] YU F,SHI X X,WANG D X,etal.Effects of insect infestation onQuercusalienavar.acuteserrataacorn dispersal in the Qinling Mountains,China[J].NewForests,2015,(46):51-61.
[23] YU F,SHI X X,WANG D X,etal.Seed predation patterns favor the regeneration of dominant species in forest gaps compared with the understory in an oak-pine mixed forest[J].ActaTheriol.,2014,(59):495-502.
[24] CHAI Z ZH(柴宗政),WANG D X(王得祥),etal.Niche characteristics of main plant populations in naturalPinustabulaeformiscommunities in Qinling Mountains,Northwest China[J].ChineseJournalofEcology(生态学杂志),2012,31(8):1 917-1 923.
[25] DU D L(杜道林),LIU Y CH(刘玉成),LI R(李 睿).Studies on the interspecific association of dominant species in a subtropicalCatanopsisfargesiiforest of Jinyun Mountain,China[J].ActaPhytoecologicaSinica(植物生态学报),1995,19(2):149-157(in Chinese).
[26] LEI R D(雷瑞德),PENG H(彭 鸿),CHEN C G(陈存根).Types and phytoenosis of natural secondary forests at Huoditang Forest Region[J].JournalofNorthwestForestryCollege(西北林学院学报),1996,11(S1):43-52(in Chinese).
[27] YANG Q(杨 琦),ZHOU J(周 婧),TAO CH(陶 楚),etal.Comparative study on interspecific association in tropical evergreenmonsoonelfinforest at two successional stages in Tongguling of Hainan Island[J].ChineseAgriculturalScienceBulletin(中国农学通报),2014,30(22):8-15(in Chinese).
[28] JU T ZH(巨天珍),KANG L D(康丽丹),WANG L F(王立峰),etal.Interspecific association analysis of dominant species in shrub layer ofPinusarmandiicommunity in Xiaolongshan Mountain of Gansu Province[J].ActaBot.Boreal.-Occident.Sin.(西 北 植 物 学 报),2010,30(6):829-834(in Chinese).
[29] WANG L(王 琳),ZHANG J T(张金屯).Interspecific association and correlation of dominant species of Lishan Mountain meadow in Shanxi Province[J].ActaBot.Boreal.-Occident.Sin.(西北植物学报),2004,24(8):1 435-1 440(in Chinese).
[30] GONG ZH W(龚直文),KANG X G(亢新刚),GU L(顾 丽),etal.Interspecific association among arbor species in two succession stages of spruce-fir conifer and broadleaved mixed forest in Changbai Mountains,northeastern China[J].JournalofBeijingForestryUniversity(北京林业大学学报),2011,33(5):29-33(in Chinese).
[31] ZHOU X Y(周先叶),WANG B S(王伯荪),etal.An analysis of interspecific associations in secondary succession forest communities in Heishiding Natural Reserve,Guangdong Province[J].ActaPhytoecologicaSinica(植物生态学报),2000,24(3):332-339(in Chinese).
[32] LEI N F(雷泞菲),SU ZH X(苏智先),etal.A comparative study on life-form spectra of evergreen broad-leaved forest in different successions in Jinyun Mountain[J].ChineseJournalofAppliedEcology(应用生态学报),2002,13(3):267-270(in Chinese).
[33] PETRITAN I C,MARZANO R,PETRITAN A M,etal.Overstory succession in a mixedQuercuspetraea-Fagussylvaticaold growth forest revealed through the spatial pattern of competition and mortality[J].ForestEcologyandManagement,2014,326:9-17.
[34] TAN Y B(谭一波),ZHAN CH A(詹潮安),YANG H D(杨海东),etal.Inter-specific associations among main tree species inMachilus chinensiscommunities in Nan’ao Island,Guangdong Province[J].JournalofCentralSouthUniversityofForestry&Technology(中南林业科技大学学报),2012,32(11):92-99(in Chinese).
[35] GUO L J(郭连金),ZHANG W H(张文辉),LIU G B(刘国彬).Species diversity and interspecific association in development sequence ofHippophaerhamnodiesplantations in loess hilly region[J].ChineseJournalofAppliedEcology(应用生态学报),2007,18(1):9-15(in Chinese).
[36] ZHAO Y H(赵永华),LEI R D(雷瑞德),HE X Y(何兴元),etal.Niche characteristics of plant populations inQuercusalienavar.acuteserratastands in Qinling Mountains[J].ChineseJournalofAppliedEcology(应用生态学报),2004,15(6):913-918(in Chinese).
[37] CAO CH(曹 春),JU T ZH(巨天珍),etal.Study on niche of main population ofQuercusalienavar.acuteserrataforest on to the Mt.Xiaolongshan in Gansu Province[J].JournalofGansuAgriculturalUniversity(甘肃农业大学学报),2006,41(3):65-69(in Chinese).
[38] SHI Z M(史作民),CHENG R M(程瑞梅),LIU SH R(刘世荣).Niche characteristics of plant populations in deciduous broad-leaved forest in Baotianman[J].ChineseJournalofAppliedEcology(应用生态学报),1999,10(3):265-269(in Chinese).
[39] ZHENG CH CH(郑超超),YI L T(伊力塔),etal.Interspecific relationship and canonical correspondence analysis of the dominant species in ecological service forest of Jiangshan City in Zhejiang Procince[J].ActaEcologicaSinica(生态学报),2015,35(22):1-11(in Chinese).
猜你喜欢
河南师范大学学报(自然科学版)(2022年5期)2022-08-08
作文周刊·小学一年级版(2022年24期)2022-06-18
绿色科技(2020年1期)2020-11-29
蔬菜(2018年5期)2018-05-17
现代农村科技(2018年3期)2018-01-18
星星·散文诗(2017年31期)2017-12-29
散文选刊·下半月(2017年7期)2017-07-31
西部林业科学(2015年6期)2016-01-27
杂草学报(2015年2期)2016-01-04
中国海洋大学学报(自然科学版)(2014年12期)2014-02-28
