
高温、加富CO2耦合对温室嫁接黄瓜光合作用日变化的影响
2016-04-25 02:17崔世茂杨志刚孙世军
北方农业学报 2016年6期
潘 璐,崔世茂,宋 阳,杨志刚,孙世军,杜 宪
(内蒙古农业大学农学院,内蒙古呼和浩特 010019)
黄瓜(Cucumis sativus L.)是设施栽培重要的蔬菜作物,喜温不耐高温,光合作用适温为25~30℃[1],但是,这是在自然条件或是某种特定的环境下测量得到的结果,随着光照、水分、肥料以及气体浓度等环境条件以及植物自身生长状况不同,其光合作用适宜温度也会发生改变[2]。光合作用是植物重要的生理生化过程,是植物生长发育的基础和生产力高低的决定性因素[3],对温度变化非常敏感,许多研究表明,植物的净光合速率随着温度的升高会有较大幅度的降低[4-6]。CO2是光合作用重要的原料,大量研究表明增施CO2可以在短期内提高植物光合效率,增加产量[7-9],同时还能够提高作物的光合最适温度[10]。另外,有研究结果显示,高温条件下加富CO2能够缓解高温胁迫,提高作物的光合能力[11-12]。但这些研究多集中在对一天之中上午的某一时间段(9:00—11:00),以及施加某一较大范围的CO2浓度(1000~1500 mol/L)进行研究,而对高温条件下全天不同时间段、施加不同浓度CO2的光合作用研究尚未见到报道。因此,本试验旨在通过研究高温与不同CO2浓度相互作用对温室嫁接黄瓜光合作用日变化的影响,探究高温、加富CO2耦合对设施黄瓜光合作用的调控机理。
1 材料和方法
1.1 供试材料
温室嫁接黄瓜“津优35号”,选用“云南黑籽南瓜”作为砧木。
1.2 试验设计
试验于2015年3—7月在内蒙古农业大学教学试验基地日光温室中进行。3月27日砧木播种育苗,3月29日接穗播种育苗,砧木和接穗均采用穴盘育苗的方法。4月15日嫁接,5月9日定植。
试验在同一温室内进行,各处理之间用塑料膜完全隔开,使其相互独立。设置5个处理,每个处理设1个小区,每个处理设3次重复。5月25日开始加富CO2。各处理具体操作如下。
高温+高浓度CO2处理:采用全封闭不放风的办法(晴天温度达到40~50℃ ,维持4~5 h),同时全天4次加富高浓度的CO2(1200~1600 mol/L)。
高温+中浓度CO2处理:采用全封闭不放风的办法(晴天温度达到40~50℃ ,维持4~5 h),同时全天4次加富中浓度的CO2(900~1200 mol/L)。
高温+低浓度CO2处理:采用全封闭不放风的办法(晴天温度达到40~50℃ ,维持4~5 h),同时全天4次加富低浓度的CO2(500~900 mol/L)。
高温处理:采用全封闭不放风的办法(晴天温度达到 40~50℃,维持 4~5 h)。
对照处理:采取常规管理模式,即每天9:00—18:00 放风。
CO2加富方法:采用乌兰察布市慧明科技有限公司生产的AⅠ型二氧化碳发生器加富CO2。
试验分别于处理7 d、14 d和21 d进行,在7:00—17:00间在内蒙古农业大学教学试验基地日光温室中进行测定。
1.3 测定项目与测定方法
1.2.1 温度、CO2浓度日变化:用北京旗硕基业科技有限责任公司生产的旗硕“农用通”,于处理7 d、14 d和21 d的7:00—17:00间进行实时测定。
1.2.2 净光合速率(Pn)、胞间 CO2浓度(Ci)、气孔导度(Gs)、蒸腾速率(Tr):用美国 LICOR 公司生产的LI-6400便携式光合仪,于上午9:00—10:00选取中间功能叶,每片叶子测定3次取平均值,共测定3个叶片,共9个数据取平均值。
1.4 数据处理
采用Microsoft Excel 2007软件对数据进行处理和作图,SAS 9.0软件进行统计分析。
2 结果与分析
2.1 不同处理温度和CO2浓度的日变化
从图1-a可以看出,不同处理温度日变化均在13:00 时达到最大值,4 个高温处理在 11:00—15:00之间的平均温度保持在40~50℃,达到了试验要求。经方差分析,与对照相比,高温处理在每天9:00以后温度达到差异显著(P<0.05)。
由图1-b可知,早晨未施加CO2之前,各处理的CO2浓度基本相同,能够达到500 mol/L左右,这是由于夜晚植物进行呼吸作用释放大量的CO2造成的。3个CO2加富处理在施加CO2之后CO2浓度均迅速上升,并在9:00之后达到各试验要求的CO2浓度;高温和对照处理的CO2浓度在7:00之后逐渐下降,15:00之后缓慢上升。
2.2 不同处理对黄瓜叶片净光合速率的影响
如图2所示,高温与不同CO2浓度耦合处理对温室黄瓜叶片净光合速率日变化有明显的影响,并且随着处理时间的推移,产生的影响也不同。处理第7天时(图2-a),不同处理黄瓜叶片净光合速率日变化总体表现为:高温+高浓度CO2>高温+中浓度CO2>高温+低浓度CO2>对照>高温,并且高温+高浓度CO2和高温+中浓度CO2在11:00左右达到最大值,高温+低浓度CO2、对照及高温在9:00左右达到最大值,说明初期施加高中浓度CO2能够提高黄瓜叶片净光合速率,并缩短叶片午休的时间;处理第14天时(图2-b),不同处理黄瓜叶片净光合速率日变化总体表现为:高温+中浓度CO2>高温+高浓度CO2>高温+低浓度CO2>对照>高温,除了高温+中浓度CO2在11:00时出现最高峰,其他4个处理的净光合速率均在9:00时到达最大值;处理第21天时(图2-c),不同处理黄瓜叶片净光合速率日变化总体表现为:高温+中浓度CO2>高温+低浓度CO2>高温+高浓度CO2>对照>高温,各处理的最大值均出现在9:00左右。从全天的变化来看,高温+高浓度CO2、高温+中浓度 CO2、高温+低浓度 CO2及对照的黄瓜叶片净光合速率日变化在处理期间均呈双峰曲线,而高温处理呈单峰曲线。同时,从图2中可以看出,高温+高浓度CO2在处理初期的净光合速率最大,随着处理时间的延长,净光合速率显著降低,与处理第7天和第14天时相比,在21 d时的平均净光合速率分别降低了90.78%和65.68%;高温+中浓度CO2、高温+低浓度CO2和对照随着处理时间的推移,平均净光合速率呈先增加后降低的趋势,21 d后均显著降低;高温处理在处理期间的净光合速率均最小,且平均净光合速率随着处理时间的增加而降低(表 1)。
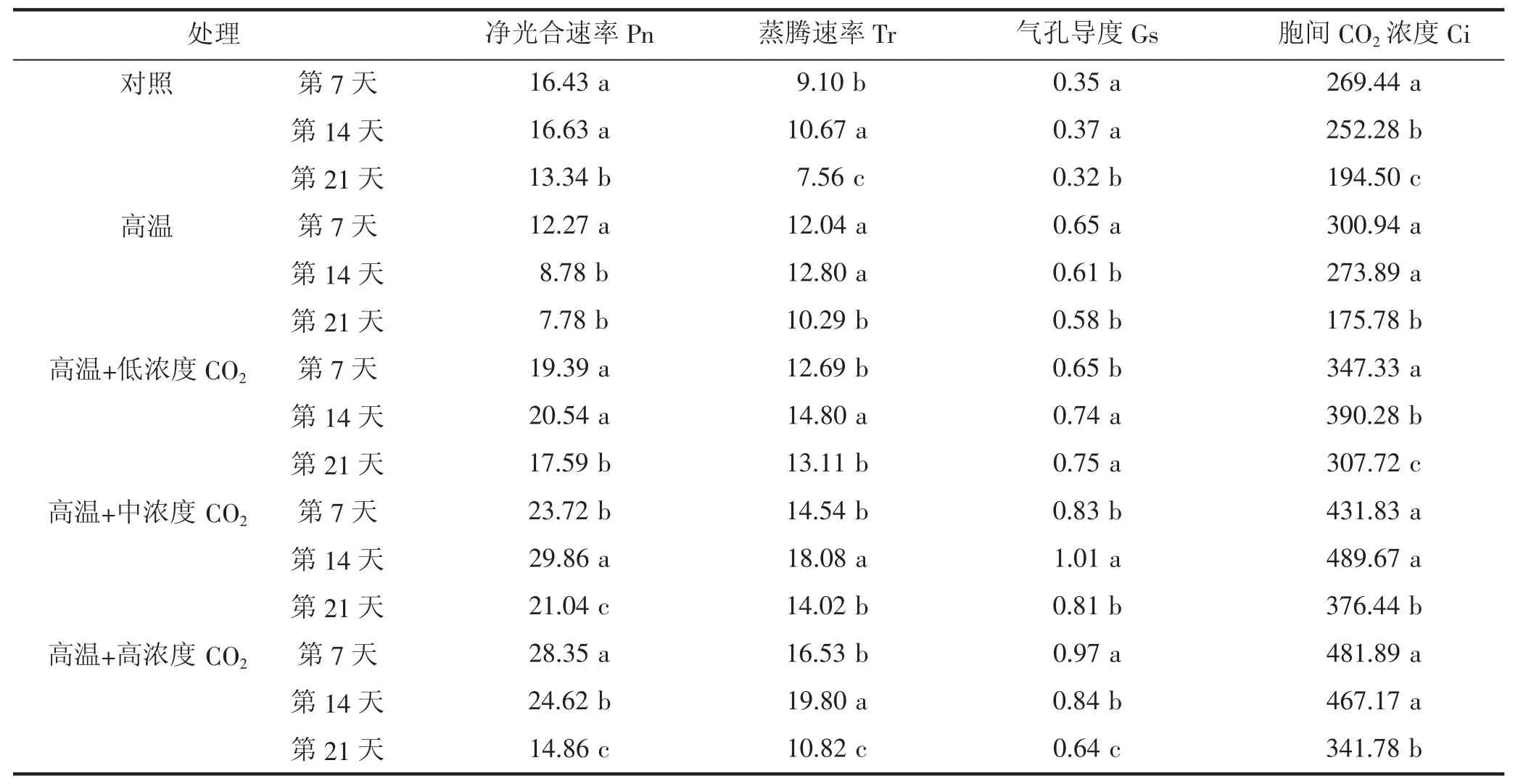
表1 不同处理黄瓜叶片净光合速率、蒸腾速率、气孔导度、胞间CO2浓度日变化平均值
2.3 不同处理对黄瓜叶片蒸腾速率的影响
由图3可知,各处理第7天和第14天,不同处理黄瓜叶片蒸腾速率日变化总体表现为:高温+高浓度 CO2>高温+中浓度 CO2>高温+低浓度 CO2≈高温>对照;而在处理第21天时,其日变化总体表现为:高温+中浓度CO2≈高温+中浓度CO2>高温+低浓度CO2≈高温>对照。无论是处理7 d、14 d还是21 d,黄瓜叶片蒸腾速率的日变化均为单峰曲线,其变化趋势与温度变化趋势一致(图1-a),即从7:00后蒸腾速率上升,到13:00达到最大值,之后呈下降趋势;并且各处理的日平均蒸腾速率均是随着处理时间的增加,先上升后下降(表1),这是由于蒸腾速率主要受到温度影响,在最适温度范围,随着温度的上升而上升,之后温度上升会导致气孔关闭,蒸腾速率降低[13]。同时,从全天来看(表1),各处理21 d后黄瓜叶片平均蒸腾速率均显著降低,说明随着植株衰老,叶片的蒸腾作用减弱。
2.4 不同处理对黄瓜叶片气孔导度的影响
从图4可以看出,不同时间的各处理黄瓜叶片气孔导度的日变化趋势与蒸腾速率日变化一致,均是呈先增高后降低的单峰曲线,并且在13:00时达到最高峰,但与蒸腾速率日变化的趋势相比,变化较为平缓。这是因为,蒸腾速率是气孔导度在植物个体水平的表现[14],植物的蒸腾速率会随着气孔导度的变化而变化。这一结果与净光合速率日变化趋势不同(图2),说明高温、加富CO2耦合处理对温室黄瓜净光合速率的影响可能不是由于气孔因素造成的,这一结果与李天来[3]等在番茄上的研究结果相同。
2.5 不同处理对黄瓜叶片胞间CO2浓度的影响
图5表明,从全天的变化来看,无论是处理第7天、第14天还是第21天,不同处理的黄瓜叶片胞间CO2浓度在一天中均呈早晚高、中午又回升的近“W”型曲线变化趋势,其变化趋势与净光合速率变化结果相反,即在早晨(7:00)和傍晚(17:00)净光合速率较弱的时候,胞间CO2浓度最大,随着净光合速率的增加,叶片固定较多的CO2,导致胞间CO2浓度下降;之后,胞间CO2浓度随着净光合速率在“午休”时减弱而上升;同时,与高温和对照相比,高温与不同浓度CO2耦合均提高了黄瓜叶片的胞间CO2浓度,但高温+高浓度CO2在处理初期(7 d)时,胞间CO2浓度日变化均高于其他处理,随着处理时间的延长,胞间CO2浓度逐渐降低,而高温+中浓度CO2与高温+低浓度CO2在处理期间的胞间CO2浓度呈先上升后下降的趋势(表1),在处理第14天后,高温+中浓度CO2的胞间CO2浓度日变化高于其他各处理。
3 讨论
随着全球大气CO2浓度升高产生的温室效应,使CO2和温度成为全球气候变化对农作物产量影响最为重要的两个因子。目前,大气CO2浓度上升以及气温升高对作物光合作用的影响成为国内外研究新的热点,以此来了解未来气候变化对作物光合生理的影响[15]。因此,目前,国内外研究已经开始将目光从高温对植物的危害转向对高温的利用,例如,温度升高使红桦器官的可溶性蛋白总量增加[16];适度增温有利于拟南芥生长,对增加叶绿素含量和叶绿体数量都有作用[17]。关于温度的利用,更多的是与CO2交互作用的研究:同时升高温度、增加CO2浓度,提高了柿树、葡萄、西葫芦、结球莴苣和水稻的净光合速率、群体最大光合速率以及加速了植株器官发育[18-19];大气CO2浓度和温度升高均能促进草本植物株高、基茎、叶片数目和生物量的增加,并可以减少草本植物的黄叶数目[20]等。王玉静、张丽红及笔者[11-12,21]等人也对高温、加富CO2条件下温室黄瓜光合作用及植株生长做了研究,结果表明,高温条件下增施CO2确实提高了黄瓜叶片的光合能力、细胞保护酶活性以及黄瓜单瓜重等。本试验的结果同样证明,与对照和高温处理相比,高温条件下无论增施怎样浓度的CO2均可以提高黄瓜叶片的净光合速率。
植物的光合作用是一个极其复杂的生理生化过程,光合作用的强弱及其日变化状态是各种内外因素综合作用的结果[3]。许多研究已经表明,植物的光合作用,在中午高温和强光作用下会出现“午休”现象,即植物的净光合速率在中午会出现明显的降低,形成净光合速率日变化的“双峰”曲线[3,22-23],这一结果与本试验结果一致。本试验结果还表明(表1、图3),高温条件加富高浓度CO2在处理前期净光合速率增加明显,且可以延长净光合速率日变化高峰出现的时间,减少“午休”时间,但随着处理时间的延长平均净光合速率降低,净光合速率日变化的高峰出现时间提前;高温条件加富中浓度CO2在整个处理期间均保持相对较高的净光合速率,并在14 d之前均可以延缓净光合速率日变化高峰出现的时间。
植物的光合作用主要受到气孔和非气孔限制,关于植物光合作用产生的“午休”现象,许多研究结果显示主要是由于蒸腾作用的影响,是气孔限制导致的。而本试验及笔者[21]先前对高温、加富CO2对温室黄瓜光合作用日变化的研究结果显示,黄瓜叶片净光合速率日变化出现“午休”现象主要受到非气孔限制,在本试验中,不同时期的高温处理的蒸腾速率均高于对照,而净光合速率却明显降低;同时先前研究结果显示,高温加富CO2可以通过增加叶片胞间CO2浓度和光合作用关键酶Rubisco的活性来提高净光合速率,因此高温、加富CO2耦合对温室黄瓜光合作用的调控主要受到非气孔因素的调节。
4 结论
在不同时期,高温、加富不同浓度CO2均可提高温室黄瓜叶片的净光合速率以及胞间CO2浓度,但高温加富高浓度CO2对光合作用的提升效果在处理前期(7 d)显著,后期效果逐渐减弱,高温加富中浓度CO2能够在整个处理期间均保持较高的净光合速率,影响光合作用的效果明显。
随着黄瓜植株的生长,不同处理的日平均净光合速率均会显著下降,这是植株衰老的正常表现,但高温、加富CO2耦合处理,特别是高温加富中浓度CO2,在整个处理期间的净光合速率均显著高于对照和高温处理。
参考文献:
[1]郑文法.黄瓜设施栽培的研究进展[J].绿色科技,2013,(7):60-62.
[2]李合生.现代植物生理学(第2版)[M].北京:高等教育出版社,2006.
[3]李天来,李 淼.短期昼间亚高温胁迫对番茄光合作用的影响[J].农业工程学报,2009 ,25(9):220-225.
[4]Mehdi L,Daniel E,Michel D.Effects of drought precondition on thermo tolerance of photosystemⅡand susceptibility of photosynthesis to heat stress in cedar seeding[J].Tree Physiology,2000,20(18):1235-1240.
[5]屠小菊,汪启明,饶力群.高温胁迫对植物生理生化的影响[J].湖南农业科学,2013(13):28-30.
[6]刘杰才,崔世茂,吴玉峰,等.CO2加富下空气湿度调控对高温大棚嫁接黄瓜逆境生理的影响[J].华北农学报,2012,27(3):130-135.
[7]Peet M M,Huber S C,Patterson D T.Acclimation to high CO2in monoecious cucumber.ⅡCarbon exchange rates.Enzyme activities and starch and nutrient concentra tion[J].Plant Physiol,1986,80:63-67.
[8]王为民,王 晨,李春俭,等.大气二氧化碳浓度升高对植物生长的影响[J].西北植物学报,2000,20(4):676-683.
[9]蒋高明,渠春梅.北京山区辽东栎林中几种木本植物光合作用对CO2浓度升高的响应[J].植物生态学报,2000,24(2):204-220.
[10]Joseph C V Vu,Leon H Allen.Growth at elevated CO2delays the adverse effects of drought stress on leaf photosynthesis of the C4 sugarcane [J].Journal of plant physiology,2008,166(2):107-116.
[11]王玉静,崔世茂,方 浩,等.CO2加富、高温处理对温室嫁接黄瓜幼苗质膜透性和保护酶活性的影响[J].华北农学报,2012,27(1):159-163.
[12]张丽红,宋 阳,张之为,等.长期增施CO2条件下黄瓜叶片淀粉积累对光合作用的影响[J].园艺学报,2015,42(7):1321-1328.
[13]于 强,任保华,王天铎,等.C3植物光合作用日变化的模拟[J].大气科学,1998,22(6):867-880.
[14]郑凤英,彭少麟.不同尺度上植物叶气孔导度对升高CO2的响应[J].生态学杂志,2003 ,22(1):26-30.
[15]林伟宏.植物光合作用对大气CO2浓度升高的反应[J].生态学报,1998,18(5):529-538.
[16]徐 燕.CO2浓度和温度升高对红桦可溶性蛋白含量和分配的影响[D].成都:中国科学院成都生物研究所,2007:41.
[17]Jin B,Wang L,Wang J,et al.The effect of experimental warming on leaf functional traits,leaf structure and leaf biochemistry in Arabidopsis thaliana [J].BMC Plant Biology,2011,11(35):3-10.
[18]谢 强,石 磊,杜 峰,等.CO2、温度对葡萄群体光合作用的影响[J].上海交通大学学报,2007,25(2):110-114.
[19]李萍萍,胡永光,赵玉国,等.增施CO2气肥对温室结球莴苣光合作用影响的综合模型[J].农业工程学报,2001,17(3):70-75.
[20]石 冰,王开运,邹春静,等.大气CO2浓度和温度升高对草本植物生长的影响[J].现代农业科技,2008(15):15-16.
[21]潘 璐,刘杰才,李晓静,等.高温和加富CO2温室中黄瓜Rubisco活化酶与光合作用的关系[J].园艺学报,2014,41(8):1591-1600.
[22]陈兆波,张 翼,王 沛,等.香紫苏开花期蒸腾和光合作用日变化特征及其影响因子研究[J].西北植物学报,2007,27(6):1202-1208.
[23]陈铁柱,蒋舜媛,孙 辉,等.羌活光合作用日变化及其与生理生态因子的关系[J].西北植物学报,2010,30(3):557-563。
猜你喜欢
农业工程学报(2022年4期)2022-04-24
西南农业学报(2021年7期)2021-08-23
节水灌溉(2021年5期)2021-05-26
农业与技术(2020年12期)2020-07-04
新课程·下旬(2017年7期)2017-08-14
Coco薇(2016年5期)2016-06-03
考试周刊(2016年6期)2016-03-11
少儿科学周刊·儿童版(2015年10期)2015-11-07
江苏农业科学(2015年8期)2015-09-10
湖北农业科学(2014年13期)2014-08-28
