
不同种植方式对水稻颖花穗部分布的影响
2016-05-30 00:53沈建凯谢振宇贺治洲林秋云尹明易俊良
热带作物学报 2016年2期
沈建凯 谢振宇 贺治洲 林秋云 尹明 易俊良


摘 要 为研究在不同种植方式下水稻穗颖花量的变化特征,设置4种不同田间种植方式,研究水稻稻穗颖花特征。试验发现:(1)稻穗颖花由穗基部向穗顶方向在稻穗呈抛物线型分布,穗中部颖花量最大;(2)随着单株水稻稻田生态空间增加,稻穗颖花总量及穗上、中、下部颖花量分别增加;稻穗一次枝梗数及其颖花量无明显增加;稻穗二次枝梗数及其颖花量和稻穗中、下部二次枝梗数增加及稻穗上、中、下部二次枝梗颖花量显著增加,其中杂交稻表现穗中、下部二次枝梗数增加,常规稻仅为穗中部二次枝梗数增加;(3)单株水稻颖花量与其籽粒重显著正相关(R2=0.804)。研究结果表明不同种植方式对水稻颖花穗部分布影响主要表现在二次枝梗数总数及分布在穗中、下部位的二次枝梗数,和二次枝梗的颖花量及其在穗的下、中、上部位的分布数量。
关键词 栽培生态;宽窄行;穗库容量;一次枝梗;二次枝梗
中图分类号 S314 文献标识码 A
The Effect of Different Rice Planting Patterns on
Spikelet of Rice Panicle
SHEN Jiankai1*, XIE Zhenyu1, HE Zhizhou1, LIN Qiuyun1, YIN Ming1, YI Junliang2
1 Tropical Crops Genetic Resources Institute, Chinese Academy of Tropical Agricultural Sciences, Danzhou, Hainan 571737, China.
2 Agricultural Committee of Yongzhou, Yongzhou, Hunan 425000, China
Abstract Four different rice planting patters were carried out in paddy field of Rice Experiment Base in Danzhou City of Hainan to reveal the changing characteristics of rice spikelets in different planting patters. The results showed that: (1)The quantity of rice spikelets was a parabola on rice panicle from rice panicle basal part to rice panicle top part. (2)With the field ecological space of individual rice plants increasing, the total spikelets, top spikelets, middle spikelets and basal part spikelets of rice panicle were increased, and the number of primary branches and their spikelets were not increased significantly, and the number of secondary branches and their spikelets were increased. The number of secondary branches of hybrid rice was increased by middle and basal part secondary branches, and the number of secondary branches of inbred rice was increased by middle part secondary branches only. (3)The quantity of rice spikelets per palnt was significantly positive correlated with grain weight (R2=0.804). Based on the study results, the distribution of the spikelets of rice panicle in different planting patterns mainly showed that the total number of secondary branches and their distribution in the middle and basal part of rice panicle, and the number of spikelets of secondary branches and their distribution in different parts of the rice panicle.
Key words Planting ecology; Wide-narrow row; Sink capacity; Rice primary branches; Rice secondary branches
doi 10.3969/j.issn.1000-2561.2016.02.010
中国是人口大国,也是水稻生产和消费大国,提高水稻产量是水稻科研的永恒主题,这也是中国人多地少的国情决定的必然选择[1]。水稻产量可分解为库容量及库充实度,库容量受颖花量和粒重决定。高的库容是水稻高产的基础[2-3],为了提高水稻产量,育种工作者提出了大穗模式[4-6]、重穗模式[7]、 功能叶长[8]模式和后期功能型[9]模式等;栽培方面以扩大水稻库容、强源和库源协调已是栽培研究者的共识[10-11]。由此可见,增加水稻库容量成为提高水稻产量重要途径之一。
杨惠杰等[11]和艾治勇等[12]研究认为,在稳定穗数基础上培育大穗,通过大穗实现增加水稻库容量提高水稻产量,而吴文革等[13]研究认为增加穗数并不能扩增群体的颖花量,增加穗颖花量是增加水稻库容的重要路径。两者研究结果虽然不同,但有一个共同点,就是注意到了提高水稻穗库容的必要性。水稻宽窄行栽培可优化单株水稻田间生态,水稻单穗的着粒数和籽粒的充实度增加,实现水稻高产[14-15],但并没有研究该栽培技术下对水稻是如何实现穗的库容量的增加,即颖花量的增加。为此,研究不同种植方式下的水稻穗部颖花变化及其在穗部分布变化,旨在探索这种栽培方式水稻穗颖花量增加主要是由穗部那些性状的变化而引起。
1 材料与方法
1.1 材料
1.1.1 试验地概况 试验地位于地属热带气候的海南省儋州市中国热带农业科学院热带作物品种资源研究所水稻综合试验基地。试验地土壤有机质含量30.34%,碱解氮91.17 mg/kg,速效磷44.72 mg/kg,速效钾139.17 mg/kg,pH为5.75。
1.1.2 试验材料 水稻品种杂交水稻红泰优996和常规水稻热香2号,均来自中国热带农业科学院热带作物品种资源研究所。红泰优996由红莲型不育系珞红3A为母本与RT99配组而成的优质三系杂交水稻组合。株高94.8~121.9 cm,穗长20.4 cm,每穗粒数165粒左右,结实率82%,千粒重27.4 g。株型适中,中等穗,籽粒大,穗粒结构协调,后期熟色好,产8 250~8 700 kg/hm2。热香2号为常规稻品种,株高95~105 cm左右,穗长25.6 cm,每穗粒数138粒左右,结实率89%,千粒重22.1 g,株型紧凑,分蘖力较好,平均产6 000~7 200 kg/hm2。
复合肥为富岛牌复合肥(26︰8︰19),GB21633-2008标准,海南富岛复合肥股份有限公司生产。尿素含氮量46.0%,磷肥含磷量16.0%,钾肥含钾量50%。
1.2 方法
1.2.1 试验设计 试验设置4个处理,3次重复,随机区组试验设计,小区面积6 m2,长方形小区。
处理1(T1):水稻种植规格20 cm×20 cm,株行距均为20 cm,单株水稻占地面积400 cm2,密度24万株/hm2。处理2(T2):水稻种植规格15 cm×(40+30)cm,株距15 cm,宽窄行相邻排列,宽行距40 cm,窄行距30 cm,单株水稻占地面积525 cm2,密度18万株/hm2。处理3(T3):水稻种植规格15 cm×(50+30)cm,株距15 cm,宽窄行相邻排列,宽行距50 cm,窄行距30 cm,单株水稻占地面积600 cm2,密度16.5万株/hm2。处理4(T4)水稻种植规格15 cm×(60+30)cm,株距15 cm,宽窄行相邻排列,宽行距60 cm,窄行距30 cm,单株水稻占地面积675 cm2,密度14.8万株/hm2。
1.2.2 田间管理 大田肥料N、P2O5、K2O用量按2︰1︰2计算氮肥、磷肥、钾肥的用量,其中纯氮(N)用量每亩12.0 kg。按此肥料用量红泰优996和热香2号均可以发挥出高产潜力。肥料总用量按比例5︰3︰2分为基肥、穗肥和粒肥施用。施肥时先按照复合肥的含氮量计算出每次施肥的复合肥量,磷和钾不足使用单质磷肥和钾肥补足量。除底肥采用机械混施外,其它均为人工撒施方式施肥。
水稻分蘖末期没有晒田,杂草人工拔除,其它田间管理同水稻常规生产管理。
1.3 数据采集方法及分析
水稻成熟期田间调查株高、有效穗;品字形3点取样,每点5株收割15株(穴)人工脱粒晒干后称重,作为株(穴)产量。每个处理5点取样选取长势良好,没有受到病虫草害危害的水稻5株(穴)进行室内考种,并进行穗部性状数据采集。考种时从近稻穗基部开始,依次按照一次枝梗在稻穗由穗基部向穗顶的排列,记录稻穗一次枝梗数及其颖花数、二次枝梗数及其颖花数。
按稻穗一次枝梗由穗基部到穗顶的排列,把靠近穗基部的一次枝梗定义为稻穗下部枝梗,其颖花为穗下部颖花,对应的二次枝梗及其颖花为穗下部二次枝梗和颖花;以此类推,上部枝梗和颖花量为靠近稻穗顶部的一次枝梗和及其颖花;中部枝梗和颖花即为处在稻穗中部的一次枝梗及其颖花。同理,一次枝梗上部和下部对应的二次枝梗及其颖花为穗下部和上部二次枝梗及其颖花。
数据统计和绘图使用Excel 2007,方差分析使用DPS v7.05,多重比较采用LSD法。
2 结果与分析
2.1 不同处理水稻颖花穗部分布
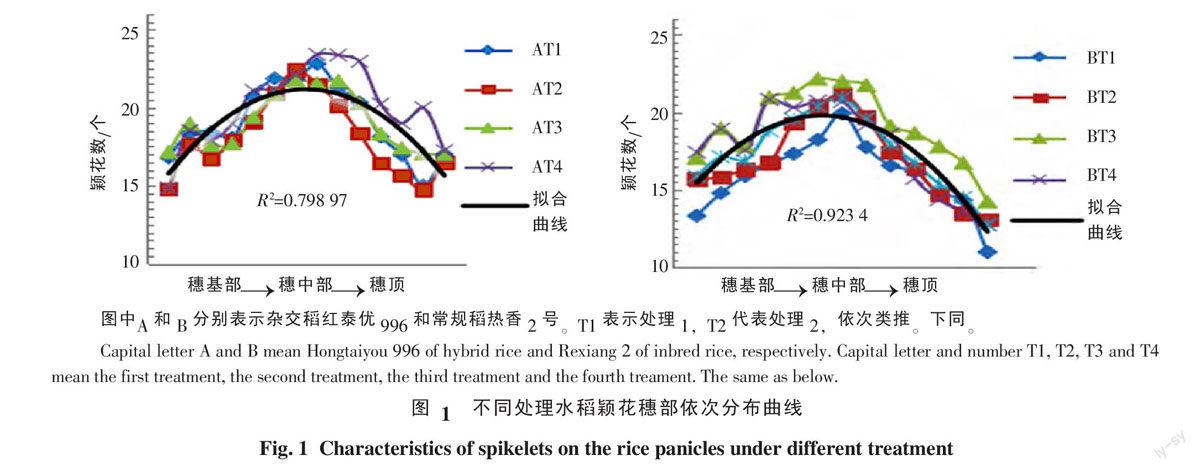
2.1.1 颖花穗部依次分布曲线 依据水稻稻穗一次枝梗稻穗上依次分布,由穗基部向穗顶依次统计一次枝梗颖花量和其对应的二次枝梗颖花量并作图(图1)发现,不论在上述哪种试验条件下,颖花在稻穗分布拟合曲线均呈现抛物线型,稻穗中部颖花量最大,穗下部颖花量和穗上部颖花量处于抛物线峰的两侧,穗下部和穗上部颖花量相对高低没有明显规律。杂交稻和常规稻相比,颖花穗部分布曲线趋势有差异。
2.1.2 颖花穗不同部位分布比较 进一步对不同种植方式下的稻穗颖花量及其在稻穗下、中、上部的分布(图2)统计分析发现,不同处理对穗颖花量有较大的影响,随着单株水稻所占稻田生态空间增加,穗总颖花量以及下、中、上部位颖花分布量均呈增加趋势。杂交稻品种AT4穗总颖花量以及穗下、上部位颖花量显著高于AT1、AT2和AT3,穗中部颖花AT4显著大于AT1和AT2,与AT3差异不显著。常规稻品种BT4和BT3穗总颖花量以及中部位颖花量显著大于BT1和BT2,BT4和BT3差异不显著;BT4、BT3穗下、上部颖花量显著高于BT1,但与BT2差异不显著。说明改变稻田微生态环境,会引起水稻稻穗颖花总量及下、中、上部颖花量发生显著变化,不同类型品种间也存在差异。
2.2 不同处理稻穗一次枝梗颖花量
2.2.1 一次枝梗数 图3可以看出,杂交稻品种和常规稻品种稻穗一次枝梗数AT4、BT4最大,处理间差异不显著。说明不同种植方式不会影响水稻一次枝梗数发生显著变化。
2.2.2 一次枝梗颖花数 图4表明,杂交稻一次枝梗总颖花量及穗下、中、上部颖花量均随着单株拥有稻田空间增加而增加,除穗下部颖花量AT4显著高于AT1外,其它处理间均差异均不显著。常规稻一次枝梗总颖花量及穗下、中、上部颖花量均随着单株稻田生态空间增加而略有减少,但差异不明显。说明水稻一次枝梗颖花量各处理间变化较小,差异不显著,即不同种植方式对一次枝梗颖花量影响较小或没有影响。
2.3 不同处理稻穗二次枝梗颖花量
2.3.1 二次枝梗数 图5可以看出,水稻二次枝梗总数及穗下、中、上二次枝梗数均表现随着单株生态空间增加而增加,二次枝梗数的增加主要表现穗中或下部二次枝梗数的增加;穗上部二次枝梗数不同处理间没有显著差异。就杂交稻品种而言,二次枝梗数增加主要来自穗下部和中部枝梗数的增加。就常规稻品种而言,二次枝梗数的增加主要来自穗中部二次枝梗数的增加。
2.3.2 二次枝梗颖花量 图6可以看出,水稻穗二次枝梗颖花总量和穗下部、中部、上部颖花量均随着单株所占空间的增大而增加。杂交稻二次枝梗颖花总数和穗上部颖花总数AT4显著大于AT1、AT2和AT3;穗下部颖花量AT4显著高于AT3并高于AT1和AT2;穗中部颖花量AT4显著大于AT1和AT2,与AT3差异不明显。常规稻二次枝梗颖花总量、穗下部颖花量和穗中部颖花量均表现BT4显著大于BT1和BT2,与BT3差异不显著,穗上部颖花量BT4、BT3和BT2间差异不显著,但显著大于BT1。说明单株水稻稻田生态空间增加导致水稻二次枝梗颖花总量及其穗上、中、下部颖花量显著增加。
2.4 不同处理水稻颖花与产量的相关性
由图7可以看出,水稻单株颖花量与单株产量呈正相关,说明水稻单株颖花量的增加是其产量的增加主要原因。
3 讨论与结论
穗是水稻的最重要器官,承载着水稻产量的最终表现形式,穗部性状的变化也最能反映出水稻受到外部生态环境变化影响的程度。稻穗枝梗数和颖花量受到品种基因型、生态环境条件和栽培技术等多方面因素的影响,有明显基因型和环境效应[16-17]。包达[18]研究发现在不同地点、不同年份超级稻产量及产量结构表现多样性变化,产量越高,产量构成要素协调性越好;张卫星等[19]研究发现水稻干旱胁迫下稻穗二次枝梗数显著减少,穗颖花量明显下降;郑华斌等[20]研究发现水稻垄栽有效穗和穗粒数都显著增加。笔者发现不同株行配置条件下水稻单株颖花量差异显著,单株水稻稻田生态空间增加,稻穗颖花量显著增加,分布在稻穗下、中、上部的颖花量均表现显著增加。研究也发现,单株水稻颖花量与其籽粒重表现正相关(R2=0.804),说明水稻颖花量增加是水稻产量增加的主要因素,这和吴文革等[13]和董桂春等[23]研究结果一致。
吴文革等[13]研究认为增加水稻一次枝梗及其颖花量是提高水稻颖花量的次要途径。笔者将稻穗按其一次枝梗在稻穗上分布,分为下、中、上三部分分别分析枝梗数及其颖花量特征发现,不同处理间稻穗一次枝梗数、一次枝梗总颖花量和一次枝梗下、中、上部颖花量并没有明显变化,说明稻穗一次枝梗及其颖花量不是导致不同处理水稻稻穗颖花变化的原因。分析稻穗二次枝梗及其颖花在稻穗上分布发现,稻穗二次枝梗总数、穗中或下部二次枝梗数出现显著变化,二次枝梗颖花总量及穗上、中、下部颖花量也发生明显变化,说明稻穗中或下部二次枝梗数增加和穗上、中、下部颖花量增加是水稻穗部颖花量增加的共同原因。吴文革等[13]和董桂春等[23]认为水稻穗粒数增加主要途径是增加二次枝梗数及其颖花量,与本研究结果相似。但研究也发现,随着单株水稻稻田生态空间的增加,杂交稻表现为穗中、下部二次枝梗数均增加,常规稻仅穗中部二次枝梗数增加,说明常规稻和杂交稻二次枝梗颖花量增加路径存在差异,但试验材料仅含1个杂交稻和1个常规稻,具有一定的局限性,需要进一步研究验证。
关于穗颖花(穗粒数)与产量研究发现产量与二次枝梗数、穗颖花数显著正相关[24-25],与一次枝梗数显著[25]或不显著[24]正相关。董桂春等[23]认为在保持适宜一次枝梗数的基础上,增加二次枝梗数,一次枝梗数和二次枝梗数保持合适的比例,实现每穗粒数的增加,进而提高库水稻容量[23]。沈建凯等[26]研究直播稻也发现,水稻高产二次枝梗数增加是主因之一。由此说明,二次枝梗数及其颖花量增加在提高产量方面的重要性。二次枝梗数和二次枝梗籽粒数增加是水稻大穗的生物学基础[21]。因此,水稻栽培和育种上加以对这些穗部性状的调控和选择与培育有可能实现穗大粒多,水稻库容量增加,实现水稻品种产量潜力提高。
参考文献
[1] 闵绍楷, 程式华, 朱旭东,等. 提高杂交水稻产量潜力的育种策略[C]// 中国农学会等编. 21世纪水稻遗传育种展望-水稻遗传育种国际讨论会文集. 北京: 中国农业科技出版社, 1999: 40-45.
[2] 杨惠杰, 杨仁崔, 李义珍, 等.水稻超高产品种的产量潜力及产量构成因素分析[J]. 福建农业学报, 2000, 15(3): 1-8.
[3] 黄育民, 陈启锋,李义珍. 我国水稻品种改良过程库源特征的变化[J]. 福建农业大学学报, 1998, 25(3): 271-278.
[4] 杨仁崔, 杨惠杰. 国际水稻研究所新株型稻研究进展.杂交水稻[J], 1998, 13(5): 29-31.
[5] 杨守仁, 张龙步, 陈温福, 等. 水稻超高产育种理论和方法[J]. 中国水稻科学, 1996, 10(2): 115-120.
[6] 黄耀祥, 林青山. 水稻超高产、特优质株型模式的构想和育种实践[J]. 广东农业科学, 1994(4): 1-6.
[7] 周开达, 汪旭东, 李仕贵,等. 亚种间重穗型杂交稻研究[J]. 中国农业科学, 1997, 30(5): 91-93.
[8] 袁隆平. 杂交水稻超高产育种[J]. 杂交水稻, 1997, 12(6): 1-3.
[9] 程式华, 曹立勇, 陈深广,等. 后期功能型超级杂交稻的概念及生物学意义[J]. 中国水稻科学, 2005(19): 280-284.
[10] 张洪程, 严宏生, 苏祖芳,等. 水稻高效栽培株型的研究[C]//凌启鸿主编. 稻麦研究新进展. 南京: 东南大学出版社, 1999: 99-106.
[11] 杨惠杰, 杨仁崔, 李义珍, 等. 水稻超高产的决定因素[J]. 福建农业学报, 2002, 17(4): 199-203.
[12] 艾治勇, 马国辉,青先国. 超级杂交稻生理生态特性及高产稳产栽培调控的研究进展[J]. 中国水稻科学, 2011, 25(5): 553-560.
[13] 吴文革, 张洪程, 吴桂成,等. 超级稻群体籽粒库容特征的初步研究[J]. 中国农业科学, 2007, 40(2): 250-257.
[14] 郭守斌, 帅国元,刘志贤. 水稻边际效应型高产栽培技术探讨[J]. 杂交水稻, 2008, 23(6): 40-43.
[15] 敖和军, 方远祥, 熊昌明,等. 株行距配置对超级杂交稻产量及群体光能利用的影响[J]. 作物研究, 2008, 22(4): 263-269.
[16] 姚友礼, 王余龙, 蔡建中. 水稻大穗形成机理的研究Ⅰ.品种间每穗颖花分化数的差异及其与穗部性状的关系[J]. 江苏农学院学报, 1994, 15(2): 33-38.
[17] 杨洪建, 王余龙, 黄建晔, 等. 开放式CO2浓度增高对水稻颖花分化和退化的影响[J]. 应用生态学报, 2002, 13(10): 1 215-1 218.
[18] 包 达. 控制超级稻的无效分蘖对成穗率及产量形成的影响[D]. 长沙: 湖南农业大学, 2013.
[19] 张卫星, 朱德峰, 林贤青,等. 干旱胁迫对不同穗型超级稻品种产量及穗部性状的影响[J]. 干旱地区农业研究, 2010, 28(1): 255-260.
[20] 郑华斌, 刘建霞, 姚 林,等. 垄作梯式生态稻作对水稻光合生理特性及产量的影响[J]. 应用生态学报, 2014, 25(9): 2 598-2604.
[21] 马 均,周开达. 亚种间重穗型杂交稻穗颈维管束与穗部性状的关系[J]. 西南农业学报, 2001, 14(3): 1-5.
[22] 祁玉良, 石守设, 鲁伟林,等. 不同栽植密度杂交稻分蘖成穗规律及其穗部性状研究[J]. 中国农学通报, 2006, 22(5): 177-181.
[23] 董桂春, 李进前, 董燕萍,等. 产量构成因素及穗部性状对籼稻品种库容的影响[J]. 中国水稻科学, 2009, 23(5): 523-528.
[24] 王昌华, 徐正进, 夏永胜.水稻穗部性状及其与产量相关分析[J]. 辽宁农业科学, 2007(3): 54-56.
[25] 王淑玲, 徐正进,周丹卉. 水稻穗部性状与产量及其构成因素之间的关系[J]. 中国农村小康科技, 2005(6): 28-29.
[26] 沈建凯, 郑华斌, 尹 明,等. 暗直播栽培对水稻穗部性状及产量的影响[J]. 作物杂志, 2014(4): 84-87.
