
不同年龄毛竹碳氮磷化学计量特征
2016-05-30 00:53刘广路范少辉郭宝华杜满义李兴军
热带作物学报 2016年2期
关键词:毛竹
刘广路 范少辉 郭宝华 杜满义 李兴军


摘 要 以毛竹分布中心区的不同年龄毛竹为研究对象,比较不同年龄毛竹林碳、氮、磷含量及其比值,探讨毛竹不同生长阶段主要养分及计量比值的变化特征,揭示毛竹生长规律,同时为“生长速率理论”提供证据。结果表明:毛竹同一器官碳、氮、磷含量和碳氮比、碳磷比、氮磷比随着毛竹年龄的增大差异达到显著水平(P<0.01),1~3年生竹叶、竹枝和竹根碳磷比和氮磷比呈降低的趋势,其后氮磷比升高,5年生氮磷比最高;1~3年生竹秆碳氮比、碳磷比和氮磷比与竹叶变化趋势相反,4~6 a竹秆碳磷比和氮磷比与竹叶变化趋势相同,反映了毛竹在不同的生长阶段碳、氮和磷含量和计量比值发生了变化,1~3年生毛竹具有较快的生长速度,竹根和竹叶氮磷比降低,随着毛竹生长速率的降低,氮磷比升高,符合“生长速率理论”。1~6 a竹叶N ∶ P分别为13.03、9.81、7.47、15.17、21.01、15.52,平均值为13.67,N ∶ P<14反映了研究区毛竹生长主要受到N元素的限制。其中,2、3年生N ∶ P低于10,对2、3年生竹施用N肥可以促进竹子的更好生长;5年生毛竹根和叶的氮磷比值最高,5 a时采伐可以降低土壤P素缺失风险。
关键词 毛竹;碳、氮、磷含量;计量比值
中图分类号 S718.55 文献标识码 A
The Carbon, Nitrogen and Phosphorus Contents of
Phyllostachys edulis with Different Ages
LIU Guanglu1, FAN Shaohui1 *, GUO Baohua1, DU Manyi2, LI Xingjun1
1 International Centre for Bamboo and Rattan / Key Laboratory of Science and Technology
of Bamboo and Rattan, State Forestry Administration, Beijing 100102, China
2 North China Foresty Experiment Center, CAF, Beijing 102300, China
Abstract Phyllostachys edulis with different ages was taken as the research objective to compare the content change of carbon, nitrogen and phosphorus of moso bamboo with different ages, to reveal the growth law, while providing an evidence for the“Growth Rate Hypothesis”(GRH). The results showed that C, N, P content and C ∶ N, C ∶ P, N ∶ P of the same organ showed significant difference with the increase of age. C ∶ N and N ∶ P of leaves, branches and roots decreased with the age increased from 1 year to 3 years, then the N ∶ P increased with the age of moso bamboo form 4 years to 6 years. Conversely, C ∶ N and N ∶ P of stems increased with the age increased from 1 year to 3 years. Meanwhile, the C ∶ P and N ∶ P trend of stems just as same as the trend of leaves, branches and roots from the 4 years to 6 years. The carbon, nitrogen and phosphorus content and its ratio change was observed for moso bamboo with different growth stages. Because the moso bamboo of the 1-3 years grew faster than that of the 4-6 years, the N ∶ P in roots and leaves of moso bamboo with 1-3 years was lower than that of the 4-6 years. The results were consistent to“GRH”. The N ∶ P of from the 1 to 6 years old was 13.03, 9.81, 7.47, 15.17, 21.01, 15.52, respectively. Its average was 13.67 and was lower than the threshold value of 14. It reflected the growth of moso bamboo in the area to be limited primarily by N element quantities.The lowest N ∶ P of roots and leaves appeared on the 2 and 3 years moso bamboo. It indicated the metabolism of the 2, 3-year old moso bamboo was stronger, its rhizome expanded activity, and its potential productivity was higher. N should be supplied adequately for the 2, 3-year old moso bamboo. The highest N ∶ P of roots and leaves appeared in the 5 years moso bamboo. It indicated the moso bamboo of the 5-year old moso bamboo had highest holding efficiency of N. The soil should lose less phosphorus element if the moso bamboo with the 5 years was cut.
Key words Phyllostachys edulis; Carbon, nitrogen and phosphorus content;Metering ratio
doi 10.3969/j.issn.1000-2561.2016.02.011
通过研究植物体主要化学元素(C、N、P等)的计量关系来揭示有机体的特性及行为与生态系统间的相互关系是生态化学计量学的重要研究内容,是联系分子、有机体水平和生态系统等不同水平的桥梁[1]。生长速率理论假说(Growth Rate Hypothesis,GRH)是生态计量学的基本理论之一,认为生长速率的改变必然造成元素组成及其比率的变化[2]。徐劲草和许新宜[3]在研究中国羊草生长率与全磷含量关系时发现生长速率与全磷含量存在正相关关系,而与N/P比负相关,符合生长速率假说的预测,但也有研究发现一些物种的生长不符合生长速率假说,如养分充足的植物[4]、在CO2浓度升高的情况下C3植物[5]、C4植物等[6]。C、N和P元素对生物的生长、发育及行为起着非常重要的作用,N/P比在决定植物群落结构与功能上是关键性指标[7]。通过研究植物C、N、P含量及其比值的关系,可以揭示植被生长速率与养分分配的关系,也可以为植被生长的限制性养分元素的确定提供参考依据。闫帮国等[8]通过对金沙江干热河谷地区植物C、N、P含量及其比值研究发现该区域植物生长受到N、P的双重限制,我国东部主要优势物种N含量和N/P比率明显偏低,而P含量和C/P比差异不显著,可能主要受到N的限制[9],我国陆地生态系统陆地植物C/P比和N/P比值均高于全球陆地生态系统的平均水平,系统更加缺磷[10]。竹子是我国一种特殊森林类型,有关竹类植物C、N、P含量及其比值的研究较少,竹种上主要集中在四季竹[11-12][Sinobambusa tootsik(sieb.)Makino]和雷竹[13-14](Phyllostachys praecox C. D. Chu et C. S. Chao cv. Prevernalis)上,通过对四季竹叶片和土壤N、P含量及计量比值的研究发现土壤N是影响四季竹生长的限制元素;通过对林地覆盖雷竹林土壤和叶片N、P化学计量特征的研究,表明雷竹林地的覆盖会显著改变雷竹林地土壤和立竹叶片的N、P化学计量特征,引起N、P养分的失衡,增强了P的限制性作用,导致竹林的退化。我国现有毛竹(Phyllostachys edulis)林面积443万hm2,是我国面积最大、分布最广的竹种,占我国竹林面积的74%,但有关毛竹C、N、P养分元素及计量特征的研究较少,多集中在养分含量变化[15]以及施肥[16]对竹子叶片或凋落物主要养分计量比值等方面。为此,开展了不同年龄毛竹C、N、P计量特征研究,一方面验证毛竹生长是否符合生长速率假说,为生长速率理论提供支撑;另一方面,为毛竹生长限制性元素诊断提供参考依据。
1 材料与方法
1.1 研究区概况
研究区位于福建永安市天宝岩国家级自然保护区(117°31′~117°33.5′E,25°55′~25°58′N),地处戴云山余脉,属中低山地貌,海拔580~1 604.8 m。研究区属于亚热带东南季风气候型,平均气温23 ℃,最低气温-11 ℃,最高气温40 ℃,全年无霜期290 d左右,平均降水量2 000 mm,≥10 ℃年积温4 520~5 800 ℃,持续225~250 d,空气相对湿度月均为80%左右。竹林主要分布在海拔800 m以下的红壤土上。研究区内森林覆盖率96.8%,主要为毛竹林,其间混生江南油杉(Keteleeria cyclolepis)、杉木(Cunninghamia lanceolata)、杨梅(Myrica rubra)、南酸枣(Choerospondias axillaris)、鹅掌楸(Liriodendron chinense)、木荷(Schima superba)等乔木树种[17-18]。
1.2 实验设计与样品采集分析
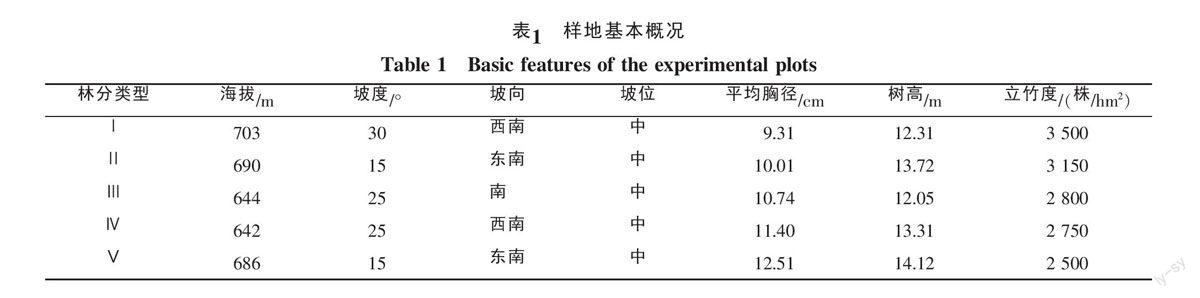
1.2.1 样地设置与调查 在对福建省天宝岩国家级自然保护区毛竹资源进行踏查的基础上,在天宝岩国家级自然保护区西洋管理所设置了不同生产力水平的毛竹林实验样地,共设置了5个生产力梯度[Ⅰ:30 000 kg/(hm2·a)、Ⅱ:34 000 kg/(hm2·a)、Ⅲ:37 000 kg/(hm2·a)、Ⅳ:42 000 kg/(hm2·a)、Ⅴ:47 000 kg/(hm2·a)],每个生产力梯度设置3块样地,共设置样地15块,样地规格为20 m×20 m。样地土壤为山地红黄壤,母岩为花岗岩,团粒状结构,质地较疏松,土层厚度65~75 cm之间,腐殖质厚度4~5 cm。对样地海拔、坡度、坡向进行记录,并调查毛竹的胸径、树高等因子[17](表1)。
1.2.2 样品采集 毛竹生物量测定采用收获法,根据样地调查资料,分别计算出不同林分类型样地中1~6年生毛竹的平均胸径,每个林分类型不同年龄的毛竹各选取2株作为标准竹砍伐,标准竹的胸径等于林分的平均胸径,每个林分类型采伐毛竹12根,共采伐毛竹60根。毛竹伐倒后,按秆、枝、叶、根、分别称取鲜重,然后各取一部分带回实验室,在60~70 ℃下烘干,以供营养元素分析。
1.2.3 样品分析 植物样品用H2SO4-H2O2消煮后,用半微量凯氏定氮法测定植物氮含量,采用钼锑抗比色法测定磷含量,碱熔火焰光度法测定钾含量[19]。
1.3 数据处理与分析
方差分析处理程序使用SPSS统计分析软件中的one way anova程序进行分析,多重比较采用LSD法进行分析;相对极差、文中图表及标准误差值为Excel所得。
2 结果与分析
2.1 毛竹碳氮磷含量及比值
2.1.1 毛竹碳氮磷平均含量变化特征 毛竹不同器官碳、氮、磷含量存在着显著差异,毛竹叶的氮、磷含量最高,竹秆的碳含量最高,不同器官之间碳、氮、磷含量差异达到极显著水平(p<0.01)(表2)。其中,碳平均含量排列顺序为竹秆>竹枝>竹叶>竹根,为431.88~528.08 g/kg,相对极差为0.20;氮平均含量排列顺序为竹叶>竹枝>竹秆>竹根,为1.49~14.18 g/kg,相对极差为0.81;磷平均含量排列顺序为竹叶>竹枝>竹秆>竹根,为1.04~0.16 g/kg,相对极差为0.73。竹秆碳平均含量最高,竹叶的氮、磷平均含量最高,竹根碳、氮、磷平均含量最低。碳、氮、磷元素平均含量在不同年龄毛竹中的变化幅度也不同,其中氮平均含量变化幅度最大,磷次之,碳变化幅度最小。
2.1.2 毛竹碳氮比、碳磷比和氮磷比变化特征
毛竹不同器官碳氮比、碳磷比和氮磷比存在着显著差异,竹叶碳氮比、碳磷比最低,竹根碳氮比最高,竹秆碳磷比最高,竹枝的氮磷比最高,不同器官的碳氮比、碳磷比、氮磷比值差异达到极显著水平(p<0.01)(表3)。其中,碳氮比的排列顺序为竹根>竹秆>竹枝>竹叶,变化范围为41.23~487.07,相对极差为1.69;碳氮比的排列顺序为竹秆>竹根>竹枝>竹叶,变化范围为488.05~3 515.30,相对极差为1.95;氮磷比的排列顺序为竹枝>竹秆>竹叶>竹根,变化范围为10.16~16.73,相对极差为0.49。竹叶氮和磷含量最高,所以碳氮比和碳磷比最低。这种结果与竹叶的生理活性最强有关,竹叶是竹子进行光合作用的场所,负责制造有机物质,而竹根和竹秆是有机物质主要的贮存器官。
2.2 不同年龄毛竹碳氮磷含量的变化特征
竹秆和竹根碳含量随着毛竹年龄的增加呈先上升后下降的趋势(图1-A)。4年生竹秆碳含量最高,平均为564.81 g/kg,4年生竹枝碳含量552.11 g/kg,4年生毛竹发育基本成熟,竹秆和竹枝的碳含量较高。竹叶的碳含量的变化趋势为1年生>2年生、3年生>4年生、5年生>6年生,这和毛竹叶片的生长周期相符合,1、3、5年生毛竹叶片均为新更新的叶片,而2、4、6年生竹叶已经生长到了第2年,其光合效率可能发生了较大变化,进而导致碳含量发了变化。竹根的变化趋势和竹叶相同,竹叶通过吸收空气中的二氧化碳和根部提供的水分和养分,在光的作用下,合成有机物质;而竹根负责提供植物生长所必须的养分和水分,竹根和竹叶碳含量具有很强的耦合作用,因而碳含量表现出相似的变化规律。
竹秆氮含量随年龄的增加呈先上升后下降的趋势(图1-B)。5年生竹秆氮含量最高,为2.63 g/kg,6年生竹秆平均氮含量最低为1.54 g/kg,竹秆中氮元素可能因为其生理活性的变化而发生转移,6年生较低的竹秆氮含量可能为合理的竹林择伐期提供数据支撑,砍伐6 a及以上竹林,氮素损失较低。1~3年生竹叶和竹枝氮含量均随着年龄的增加呈降低的趋势,4~6年生竹叶和竹枝氮含量的变化趋势则相反,5年生竹枝氮含量最高,竹叶的氮含量最低。氮元素是构成生物的最主要元素之一,在植物生命活动中起着重要作用,随着毛竹年龄的增加,毛竹生理活性降低,氮元素降低。毛竹叶片为2年生,超过一定年限的毛竹枝和叶之间可能存在氮元素的转移。除1年生竹根具有较高的氮含量外,竹根氮含量随着年龄的增加呈上升趋势,与竹秆氮含量的变化趋势相反,竹根和竹秆氮含量可能存在某种互耦关系。
竹秆磷含量随年龄的增加呈先上升后下降的趋势(图1-C)。2年生竹秆磷含量达到最高,为0.27 g/kg,其后逐步下降,表明竹秆进行的生理活动强度随年龄的增长呈降低的趋势。同时,竹秆中磷含量低于枝叶等器官的磷含量,枝叶的生理活性强度高于竹秆的生理强度。1年生竹枝和竹叶磷含量最高,反映了植物幼嫩组织具有较高的生理活性,各种生物化学反应剧烈。竹叶磷含量表现出2年生>3年生、4年生>5年生的规律,这与竹叶的生长周期相关,反映了新生竹叶具有较高的生理活性,磷含量较高。
2.3 不同年龄毛竹碳氮磷计量参数变化特征
碳氮比一方面反映了单位氮素固定碳的效率,另一方面反映了不同类型器官的分化和生长情况。1年生竹秆碳具有较高的碳氮比,竹叶、竹枝、竹根具有较低的碳氮比。毛竹在第1年的生长中快速完成从出土到秆形生长结束,在此过程中,毛竹分配更多的碳到竹秆中,而较少的碳分配到竹枝、竹根中。2~6年生竹秆碳氮比的变化趋势为先降低后上升的趋势,反映了6年生毛竹较大的固碳效率,竹枝、竹叶和竹根碳氮比的变化趋势为先升高后降低的趋势,但是其达到最高点的年龄不同,4年生竹枝的碳氮比最高,而竹叶和竹根在3年生时达到碳氮比的最高值,反映了竹根和竹叶的碳氮比具有较强的耦合性(图2-A)。
碳磷比主要反映了磷素含量与碳素之间的关系,与植物光合作用密切相关,较高的碳磷比反映了单位磷素较高的固碳效率。5年生竹秆、竹叶、竹枝、竹根的碳磷比较高,反映了5年生毛竹单位磷素的固碳效率最高,对超过5年生的毛竹进行择伐,可以降低磷素损失。竹叶和竹根的碳磷比也存在着很大的关联性,均随着年龄的增加呈先升高后降低的趋势(图2-B)。
氮磷比表示植物体同时消耗氮和磷,使氮和磷以同样的方式变化,在稳定的系统中氮磷比值保持恒定,但是在个体发育过程中,氮磷比值可能表现出某种规律性变化。5年生竹秆、竹枝、竹叶、竹根的氮磷比均为最高,分别为29.69、31.21、21.01、18.93;竹秆氮磷比随着年龄的增长呈先升高后降低的趋势,竹枝、竹叶和竹秆的氮磷比呈波浪型变化,但是不同器官的变化趋势不同,竹枝为升-降-升-降,竹叶为降-升-降,竹根为降-升-降,反映了竹根和竹叶变化趋势具有很高的同步性,而竹枝(尤其是1~4年生)氮磷比具有较大的互补性,竹秆氮磷比高,竹枝氮磷比低(图2-C)。
3 讨论与结论
毛竹叶的碳氮比、碳磷比、氮磷比分别为41.23、488.05、13.67, 与庄明浩等[20]在浙江测定的毛竹叶片碳氮比26.64、碳磷比406略高,比氮磷比15.26 略低,主要因为本研究竹叶具有较高的碳含量。竹叶碳、氮、磷含量的差异可能由区域环境的差异造成的。Reich和Oleksyn研究全球陆地植物发现,随着温度升高和年均温的降低,叶片的N和P含量增加,而N/P比降低[21],与本研究结果相符,但也有研究得出不同的结论,He等分别于2006年[22]和2008年[23]对中国草地213种优势植物的生态化学计量特征进行了研究,发现N、P含量及N/P比随着温度的变化未发生明显变化。坡度、坡向、海拔等因子也会对不同物种叶片的性质造成影响,He等[24]对中国北方草地的429份植物叶片性质进行了变异来源分析,其中环境因素可以解释变异的29%,潘复静等[25]的研究表明凋落物C值表现为上坡位较高、下坡位较低,P含量的变化规律与之相反,而N无显著变化规律,推测上坡位具有较高的N/P值;栗忠飞等[26]的研究表明处于高海拔的山地雨林幼树中C含量、C/P以及N/P比率均高于低海拔的季节雨林。本研究中毛竹叶片碳氮比、碳磷比、氮磷比与四季竹[11-12]、雷竹[14]的叶片计量比也存在一定的差异,一方面是由于物种本身的特征存在差异,另一方面可能由不同研究地点的环境的差异造成。Jackson等[27]研究全球植被细根的C ∶ N ∶ P为1 158 ∶ 24 ∶ 1,与叶片的比值相近,本研究中毛竹根的C ∶ N ∶ P为2 671 ∶ 9 ∶ 1,叶片的C ∶ N ∶ P为448 ∶ 14 ∶ 1,C/N比高于全球平均水平,毛竹根和叶具有较大的C含量和较低的N含量;N/P比低于全球的平均水平,前期的研究结果表明,毛竹林土壤主要养分计量值对毛竹林养分的计量值的影响较小[17],其差异可能源于固有的植被差异。
毛竹在不同的生长阶段碳、氮和磷元素含量和计量比值都发生了变化,1~3年生毛竹具有较快的生长速度,氮磷比降低,4~6年生竹林生长速率降低,氮磷比升高,基本符合“生长速率理论”。植物体高速率生长使更多的P分配到RNA中,进而导致生物体氮、磷元素总量发生不同速率变化,从而改变生物体内N/P[28],与本研究不同年龄氮磷含量和比值的变化趋势相符。A°gren[29]对自养生物的研究中也发现,在生长速度较低时,植物的N/P比逐渐增大,直至最大值,而后随着生长速率的增大,N/P比逐渐减小。竹根和竹叶碳氮比、碳磷比、氮磷比的变化趋势具有很大的关联性,可能反映了毛竹不同器官通过代谢进行传递的过程。植物N/P比可以反映一个地区N和P元素的相对供应水平,通过比较某个物种N/P比与区域平均N/P比的大小,可以推测植被生长所需营养元素的丰缺。Koerselman和Meuleman[30]根据在不同植物体上进行施肥实验得出,当N/P>16表示P限制,N/P<14表示N限制,但这种限制在不同物种上表现不一致。我国内蒙古地区草原的施肥实验表明,N/P>23表现为P限制,N/P<21表现为N限制[31]。本文中1~6年生竹叶N/P比分别为13.03、9.81、7.47、15.17、21.01、15.52,显示了不同年龄毛竹对N和P元素的需求不同,其中2、3年生N/P比低于10,针对2、3年生毛竹施肥可以更好促地进竹子生长。1~6年生竹叶N/P比平均值为13.67,低于14,反映了实验地所在区域毛竹的生长主要受到N的制约。植物细胞内贮藏性物质的比例随着植物物候期的推迟逐渐增多,相同植物在不同物候期下,叶片C、N、P计量特征也可能会出现很大差异[32],本研究采用在7月份一次性取样分析C、N、P元素含量值,具有一定的局限性,今后应该开展全面的、跨越整个生长季的采样,提高结论的科学性。
参考文献
[1] 贺金生, 韩兴国. 生态化学计量学: 探索从个体到生态系统的统一化理论[J]. 植物生态学报, 2010, 34(1): 2-6.
[2] Sterner R W, Elser J J. Ecological stoichiometry: the biology of elements from molecules to the biosphere[M]. Princeton University Press, 2002: 1-464.
[3] 徐劲草, 许新宜. 羊草生长率的研究和生长率假说的验证[J]. 草业科学, 2013, 30(1): 74-79.
[4] Matzek V P M , Vitousek. N: P stoichiometry and protein: RNA ratios in vascular plants: an evaluation of the growth-rate hypothesis[J]. Ecology Letters, 2009, 12(8): 765-771.
[5] Novotny A M, Schade J D, Hobbie S E, et al. Stoichiometric response of nitrogen-fixing and non-fixing dicots to manipulations of CO2, nitrogen, and diversity[J]. Oecologia, 2007, 151(4): 687-696.
[6] Polley H W, Fay P A, Jin V L, et al. CO2 enrichment increases element concentrations in grass mixtures by changing species abundances[J]. Plant Ecology, 2011, 212(6): 945-957.
[7] Tessier J T, Raynal D J. Use of nitrogen to phosphorus ratios in plant tissue as an indicator of nutrient limitation and nitrogen saturation[J]. Journal of Applied Ecology, 2003, 40(3): 523-534.
[8] 闫帮国, 何光熊, 李纪潮, 等. 金沙江干热河谷地区植物叶片中各生源要素的化学计量特征以及异速增长关系[J]. 植物生态学报, 2012, 36(11): 1 136-1 144.
[9] 任书杰, 于贵瑞, 姜春明, 等. 中国东部南北样带森林生态系统102个优势种叶片碳氮磷化学计量学统计特征[J]. 应用生态学报, 2012, 23(3): 581-586.
[10] 曾冬萍, 蒋利玲, 曾从盛, 等. 生态化学计量学特征及其应用研究进展[J]. 生态学报, 2013, 33(18): 5 484-5 492.
[11] 顾大形, 陈双林, 黄玉清. 土壤氮磷对四季竹叶片氮磷化学计量特征和叶绿素含量的影响[J]. 植物生态学报, 2011, 35(12): 1 219-1 225.
[12] 郭子武, 陈双林, 杨清平, 等. 密度对四季竹叶片C, N, P化学计量和养分重吸收特征的影响[J]. 应用生态学报, 2013, 24(4): 893-899.
[13] 刘亚迪, 范少辉, 蔡春菊, 等. 地表覆盖栽培对雷竹林凋落物养分及其化学计量特征的影响[J]. 生态学报, 2012, 32(22): 6 955-6 963.
[14] 郭子武, 陈双林, 杨清平, 等. 雷竹林土壤和叶片N, P化学计量特征对林地覆盖的响应[J]. 生态学报, 2012, 32(20): 6 361-6 368.
[15] 刘广路, 范少辉, 官凤英, 等. 不同年龄毛竹营养器官主要养分元素分布及与土壤环境的关系[J]. 林业科学研究, 2010, 23(2): 252-258.
[16] 郭子武, 虞敏之, 郑连喜, 等. 长期施用不同肥料对红哺鸡竹林叶片碳, 氮, 磷化学计量特征的影响[J]. 生态学杂志, 2011, 30(12): 2 667-2 671.
[17] 郭宝华, 刘广路, 范少辉, 等. 不同生产力水平毛竹林碳氮磷的分布格局和计量特征[J]. 林业科学, 2014, 50(6): 1-9.
[18] 杜满义, 刘广路, 范少辉, 等. 施肥对闽西毛竹生物量及碳储量分布格局的影响[J]. 热带作物学报, 2015, 36(5): 872-877.
[19] Shi L, Fan S, Jiang Z, et al. Mixed leaf litter decomposition and N, P release with a focus on Phyllostachys edulis(Carrière)J. Houz. forest in subtropical southeastern China[J]. Acta Societatis Botanicorum Poloniae, 2015, 84(2): 207-214.
[20] 庄明浩, 李迎春, 郭子武, 等. CO2浓度升高对毛竹和四季竹叶片主要养分化学计量特征的影响[J]. 植物营养与肥料学报, 2013, 19(1): 239-240.
[21] Reich P B, Oleksyn J. Global patterns of plant leaf N and P in relation to temperature and latitude[J]. Proceedings of the National Academy of Sciences of the United States of America, 2004, 101(30): 11 001-11 006.
[22] He J S, Fang J, Wang Z, et al. Stoichiometry and large-scale patterns of leaf carbon and nitrogen in the grassland biomes of China[J]. Oecologia, 2006, 149(1): 115-122.
[23] He J S, Wang L, Flynn D F, et al. Leaf nitrogen: phosphorus stoichiometry across Chinese grassland biomes[J]. Oecologia, 2008, 155(2): 301-310.
[24] He J S, Wang L, Schmid B, et al. Taxonomic identity, phylogeny, climate and soil fertility as drivers of leaf traits across Chinese grassland biomes[J]. Journal of Plant Research, 2010, 123(4): 551-561.
[25] 潘复静, 张 伟, 王克林, 等. 典型喀斯特峰丛洼地植被群落凋落物CNP生态化学计量特征[J]. 生态学报, 2011, 31(2): 335-343.
[26] 栗忠飞, 刘文胜, 张 彬, 等. 西双版纳热带雨林幼树C, N, P 的生态化学计量比对海拔变化的响应[J]. 中南林业科技大学学报, 2012, 32(5): 80-85.
[27] Jackson R B, Mooney H, Schulze E D. A global budget for fine root biomass, surface area, and nutrient contents[J]. Proceedings of the National Academy of Sciences, 1997, 94(14): 7 362-7 366.
[28] Elser J, Acharya K, Kyle M, et al. Growth rate-stoichiometry couplings in diverse biota[J]. Ecology Letters, 2003, 6(10): 936-943.
[29] A°gren G I. The C: N: P stoichiometry of autotrophs-theory and observations[J]. Ecology Letters, 2004, 7(3): 185-191.
[30] Koerselman W, Meuleman A F M. The vegetation N:P ratio: a new tool to detect the nature of nutrient limitation[J]. Journal of Applied Ecology, 1996, 33(6): 1 441-1 450.
[31] Zhang L X, Bai Y F, Han X G. Differential responses of N: P stoichiometry of Leymus chinensis and Carex korshinskyi to N additions in a steppe ecosystem in Nei Mongol[J]. Acta Botanica Sinica, 2004, 46(3): 259-270.
[32] Kerkhoff A J, Enquist B J, Elser J J, et al. Plant allometry, stoichiometry and the temperature-dependence of primary productivity[J]. Global Ecology and Biogeography, 2005, 14(6): 585-598.
猜你喜欢
江西农业(2021年24期)2021-12-07
意林·少年版(2020年13期)2020-08-02
汉语世界(2020年3期)2020-06-19
东坡赤壁诗词(2020年2期)2020-06-04
阅读与作文(小学高年级版)(2019年3期)2019-04-20
世界竹藤通讯(2018年4期)2018-09-04
浙江农林大学学报(2018年2期)2018-03-23
故事大王(2016年4期)2016-05-14
花卉(2016年17期)2016-03-07
福建农业科技(2015年1期)2015-02-27
