
不同生境紫茎泽兰构件生长及其对环境的行为响应
2016-06-27 06:08刘钊,李伟,周伟
安徽农业科学 2016年12期
刘 钊,李 伟,周 伟
(1.河西学院农业与生物技术学院,甘肃张掖 734000;2.西南林业大学林学院,云南昆明 650224)
不同生境紫茎泽兰构件生长及其对环境的行为响应
刘 钊1,2,李 伟2,周 伟2
(1.河西学院农业与生物技术学院,甘肃张掖 734000;2.西南林业大学林学院,云南昆明 650224)
摘要[目的]探究紫茎泽兰(Eupatorium adenophorum)构件生长对不同环境条件变化的响应。[方法]在哀牢山国家级自然保护区南华片区紫茎泽兰入侵的华山松(Pinus armandii)林、落叶阔叶林、林区公路边和林区空旷地等4种生境类型中共设置样方120个,测量紫茎泽兰优势植株构件数据和环境因子分析不同生境紫茎泽兰的生长状况差异性和环境因子差异性,并采用主成分分析和相关性分析研究紫茎泽兰构件生长变量主成分因子和环境变量主成分因子的关系。[结果]不同生境紫茎泽兰优势植株构件生长均差异显著(P<0.01)。在华山松林中,紫茎泽兰植株相对弱小,各枝级数分配不均,这与华山松林内的弱光照条件、高海拔的气候特征及较大的草本层盖度密切相关。林区空旷地和林区公路边的植株各枝级的分枝密度和分枝强度均较其他2种生境的大,且各枝级数分配较均匀,这与生境的强光照条件有关。落叶阔叶林中的紫茎泽兰植株粗壮茂盛,但各枝级数分配不匀,这与该生境较高的土壤肥力和夏季林内光照不足有关。[结论]环境变化是影响紫茎泽兰构件生长的关键,紫茎泽兰构件生长对环境条件变化有不同的响应。建议对保护区不同生境采取不同的管理方式,以防止紫茎泽兰的入侵或降低其入侵的危害。
关键词生境;紫茎泽兰;构件生长;环境因子;哀牢山国家级自然保护区
植物个体对环境的反应通过具有重复性的基本组成单元“构件”在数量、形状和空间排列上的变化来实现。因此,从构件水平对种群进行研究,有助于了解植物对环境的反应。因此,植物种群构件生长特性不仅是种群动态的重要方面,而且对于揭示物种与环境之间的相互作用机理具有重要意义。
目前,对紫茎泽兰(Eupatoriumadenophorum)的研究主要集中在分布、危害、生物生态学特征和防治等方面[1-3],而对种群构件生长及其对环境的行为响应则鲜见报道。Huber等[4]对薇甘菊和独叶草等构件生长的研究发现植物构件生长对环境变化有不同的响应。笔者通过比较不同生境中紫茎泽兰构件生长及环境因子的差异,揭示紫茎泽兰构件生长在不同群落中的行为响应,以期为紫茎泽兰的生态防治提供参考。
1材料与方法
1.1研究区概况哀牢山国家级保护区南华片区系原云南省大中山保护区,地处100°44′28″~100°57′42″ E,24°43′32″~25°01′10″ N,保护区总面积25 327 hm2,全年光照不足,气候垂直变化显著,从谷底到山顶海拔差1 750 m,山顶与谷底温差在10 ℃以上,该研究区具有云南亚热带北部与亚热带南部过渡区特征以及典型的山地气候特点[5]。
1.2数据收集2006年7~8月在哀牢山国家级保护区南华片区进行为期32 d的野外调查。在海拔2 200~2 500 m的范围内,紫茎泽兰主要分布于华山松林、落叶阔叶林、林区公路边和林区空旷地4种生境类型,在每种生境设置30个 5 m×5 m大小的样方,共120个生境。在样方内采集10株最高的紫茎泽兰全植株作为优势植株,测定各株的茎基粗、株高和各级分枝长度,统计各级分枝数量和各分枝上叶片数。在样方中测量海拔、坡度、坡向、乔木密度、乔木层盖度、落叶层厚度、灌木密度、灌木层盖度和土壤类型(棕黄壤、红壤、黑壤);在样方中心及四角设立5个 1 m×1 m 的小样方,调查草本植物的多度和盖度。
1.3数据处理计算不同生境紫茎泽兰各级分枝的密度、平均长度、数量、强度和分枝率。其中,分枝率以逐步分枝率表示,即为该级枝条数与下一级枝条数的比率[6]。
采用Kolmogorov Smirnov Z-检验分析不同生境紫茎泽兰构件特征因子及环境生态因子的正态性。当符合正态分布时,采用单因素方差分析;否则,采用非参数Kruskal-WallisH检验。分析不同生境紫茎泽兰构件特征及环境因子的差异,采用Duncan’s多重比较法分析不同生境间的差异显著性[7]。
对不同生境紫茎泽兰构件特征因子和环境生态因子进行标准化,然后分别进行主成分分析,比较它们分值之间的差异[7],并进行相关性分析,分析不同环境生态因子对紫茎泽兰构件生长的影响。试验数据使用Excel 2000 和 SPSS 13.0统计软件进行统计与分析。
2结果与分析
2.1不同生境的构件生长比较
2.1.1总体构件。单因素方差分析结果(F值)表明,各生境紫茎泽兰优势植株的单株茎基粗、株高、分枝数和叶片数均差异显著(P<0.01)。然而,多重比较结果表明这4个因子在两两生境间有的有差异,有的则无差异。落叶阔叶林、林区空旷地和林区公路边的紫茎泽兰各指标均显著大于华山松林(表1)。
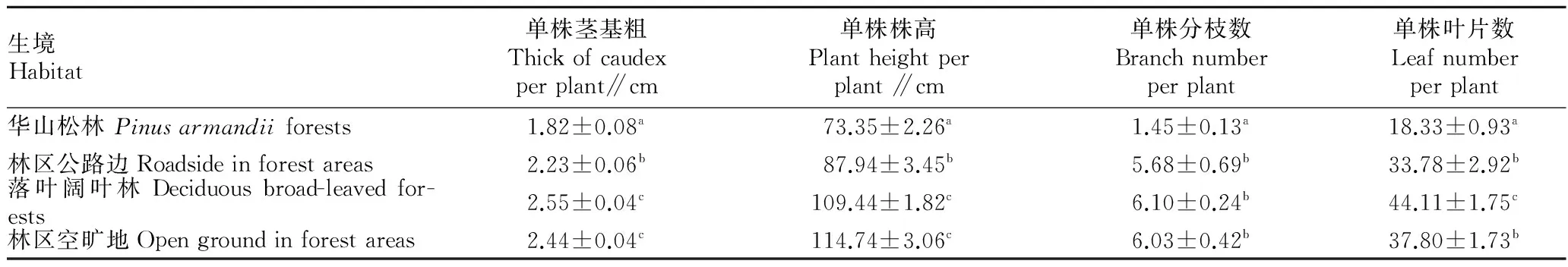
表1 不同生境紫茎泽兰构件生长情况的比较
注:同列肩标不同小写字母表示差异显著(P<0.05)。
Note:Different lowercases in the same column stand for significant difference(P<0.05).
2.1.2枝构件及比例分配。单因素方差分析(F值)及非参数 Kruskal-WallisH检验(χ2值)结果表明,分枝格局的大部分指标差异显著(P<0.01)。多重比较结果表明,各指标在两两生境间也是有的有差异,有的则无差异。华山松林中的紫茎泽兰无3级分枝;林区空旷地和林区公路边的各指标间大多差异不显著;落叶阔叶林的1级分枝各指标较大(表2)。
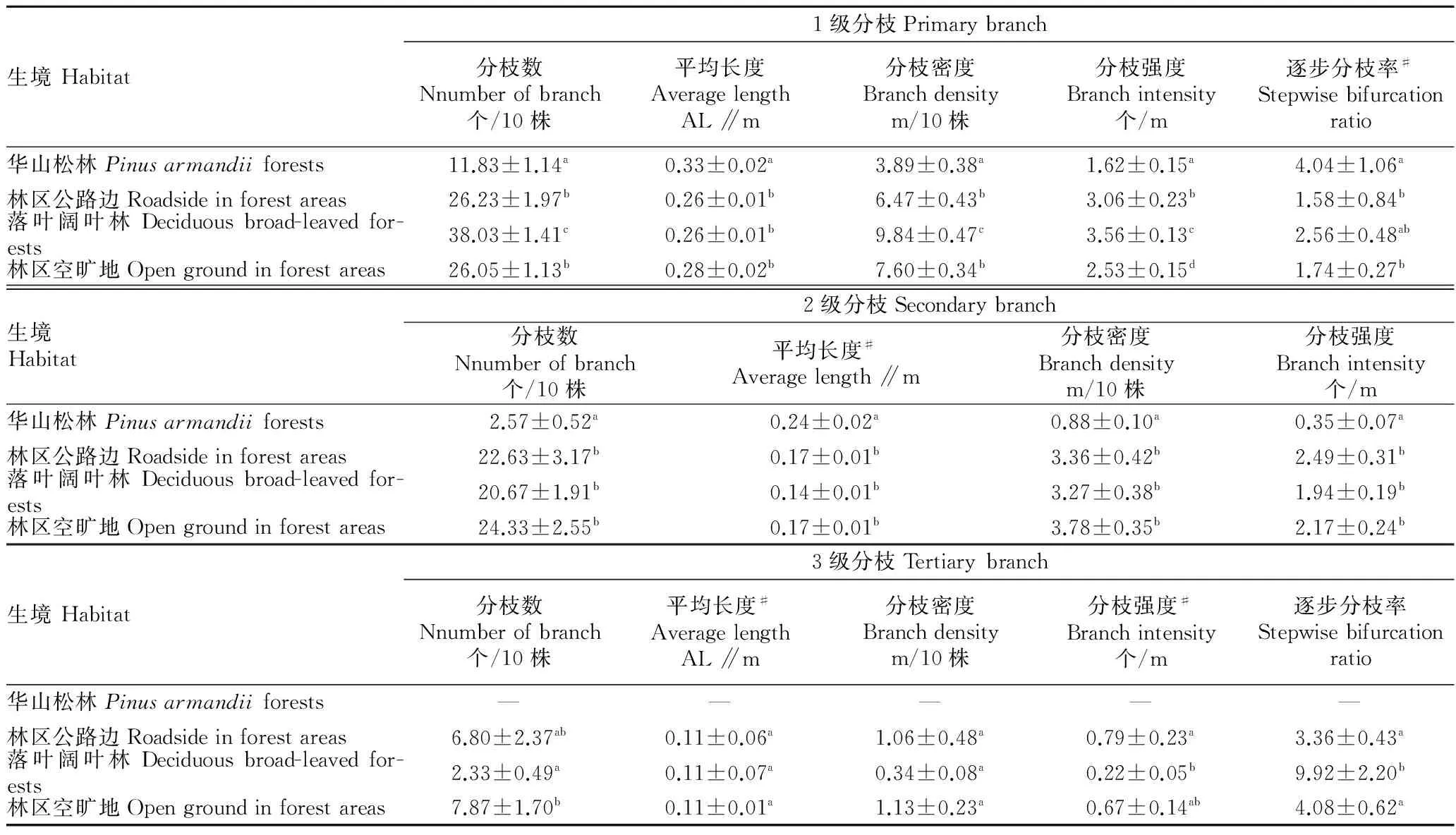
表2 不同生境紫茎泽兰枝构件的分枝格局比较
注:同列肩标不同小写字母表示差异显著(P<0.05)。#表示Kruskal-WallisH检验结果。
Note:Different lowercases in the same column stand for significant difference(P<0.05).# indicates kruskal-WallisHtest result.
从图1可以看出,不同生境紫茎泽兰1、2和3级分枝所占的比例依次减小,其中华山松林无3级分枝(图1A)。不同生境各级分枝分别在各级分枝总数中所占的比例大小不等,其中华山松林所占比例均较小;其他生境所占比例较大(图1B)。
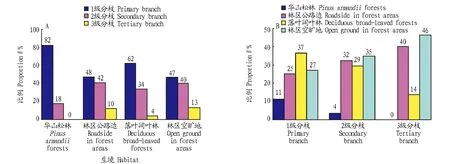
图1 不同生境各枝级分枝数的比例分配状况(A)及各枝级分枝数在各级分枝总数中的比例分配状况(B)Fig.1 The allocation in branch class level (A) and the allocation in total number of branch class level (Right) in different habitats
2.2不同生境的环境因子比较由表3可知,单因素方差分析结果表明不同生境环境因子大多存在显著差异(P<0.01)。多重比较结果表明,各指标在两两生境间同样是有的有差异,有的则无差异。
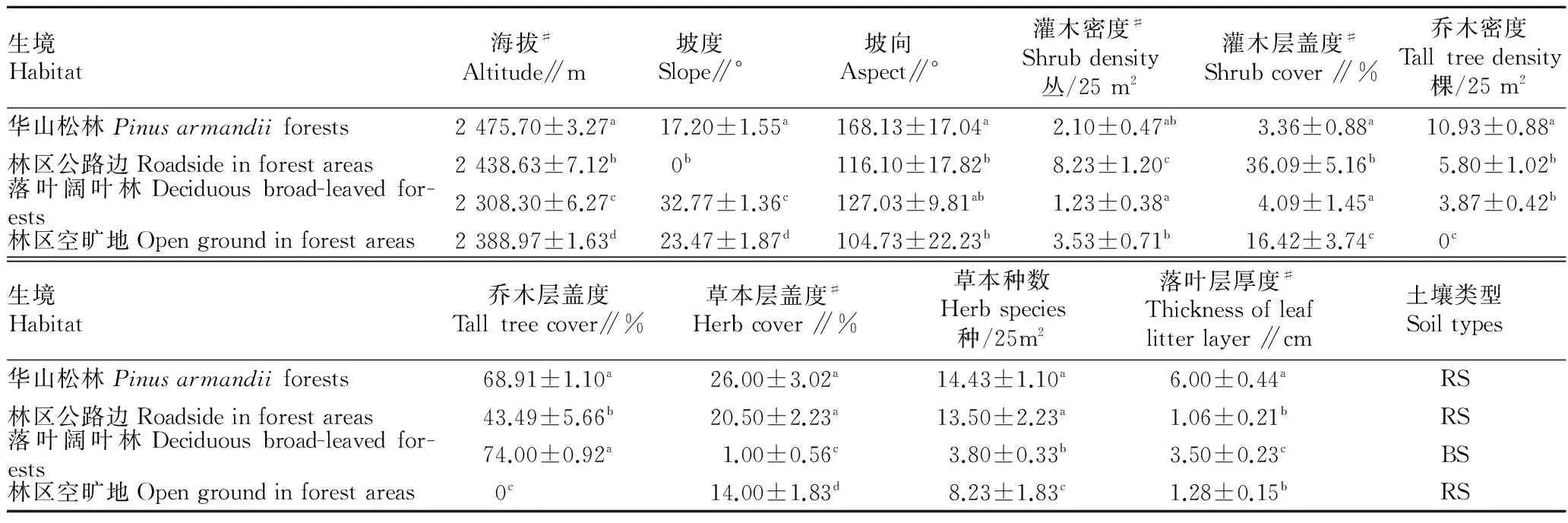
表3 紫茎泽兰不同生境的环境因子比较
注:同列肩标不同小写字母表示差异显著(P<0.05);#表示Kruskal-Wallis检验结果。
Note:Different lowercases in the same column stand for significant difference(P<0.05);#stands for Kruskal-Wallis test results.
2.3构件生长和环境因子的主成分分值比较Bartlett球形检验(P<0.01)和KMO统计量(KMO1=0.746,KMO2=0.642)均表明这些因子适合进行主成分分析。主成分分析结果表明,紫茎泽兰构件生长和环境因子的前3个主成分的累积信息量分别达72.04%和72.53%(表4)。
紫茎泽兰构件生长的主成分分值比较结果表明,第1、2主成分分值分别差异显著(F3,116=89.4,P<0.01;F3,116=10.2,P<0.01);第3主成分分值差异不显著(F3,116=2.2,P>0.05)(图2A)。环境因子主成分分值比较结果表明,第1、2和3主成分分值分别差异显著(F3,116=130.6,P<0.01;F3,116=31.4,P<0.01;F3,116=52.4,P<0.01)(图2B)。但是,就同一主成分而言,各自分布在零轴上下且偏离零轴较大的,差异显著;反之,分布在同侧或偏离零轴较小的,则差异不明显。
2.4构件生长与环境因子的关系根据各主成分涵盖的信息因子载荷系数的绝对值的大小,综合绝对值较大的因子对主成分命名。将构件生长的第1、2和3主成分依次为主要构件、分枝数量和分枝长度。将环境因子的第1、2和3主成分依次命名为地形地被因子、灌木因子和光照因子(表4)。相关性分析结果表明,主要构件与地形地被因子、光照因子呈显著负相关(P<0.01),分枝数量与灌木因子呈显著正相关(P<0.01),与光照因子呈显著负相关(P<0.01)(表5)。
3结论与讨论
3.1构件生长的行为响应该研究中大中山不同生境紫茎泽兰的构件生长具有不同的行为响应。落叶阔叶林、林区空旷地和林区公路边的总体构件生长状况显著优于华山松林,分枝密度和分枝强度也显著高于华山松林。落叶阔叶林的1级分枝密度和强度较大,但2、3级分枝的密度和强度较林区空旷地和林区公路边小。华山松林和落叶阔叶林中的紫茎泽兰各枝级间比例分配不均匀,如华山松林中没有3级分枝,落叶阔叶林的3级分枝占较小比例。林区空旷地和林区公路边的各枝级分配相对均匀,1、2和3级分枝数在各级分枝总数中占较大比例。因为固着生长个体适应环境变化的方式主要是构型随环境发生改变,产生形态可塑性,从而直接影响植株对光、空间等资源的利用和适应对策[8]。不同生境紫茎泽兰构件生长的差别较大,是构件种群通过改变形态和生物量分配,对环境条件变化做出的反应。此外,作为一个整体,在不同的环境下植物体某一性状的改变可能会引起其他性状的变化[6]。
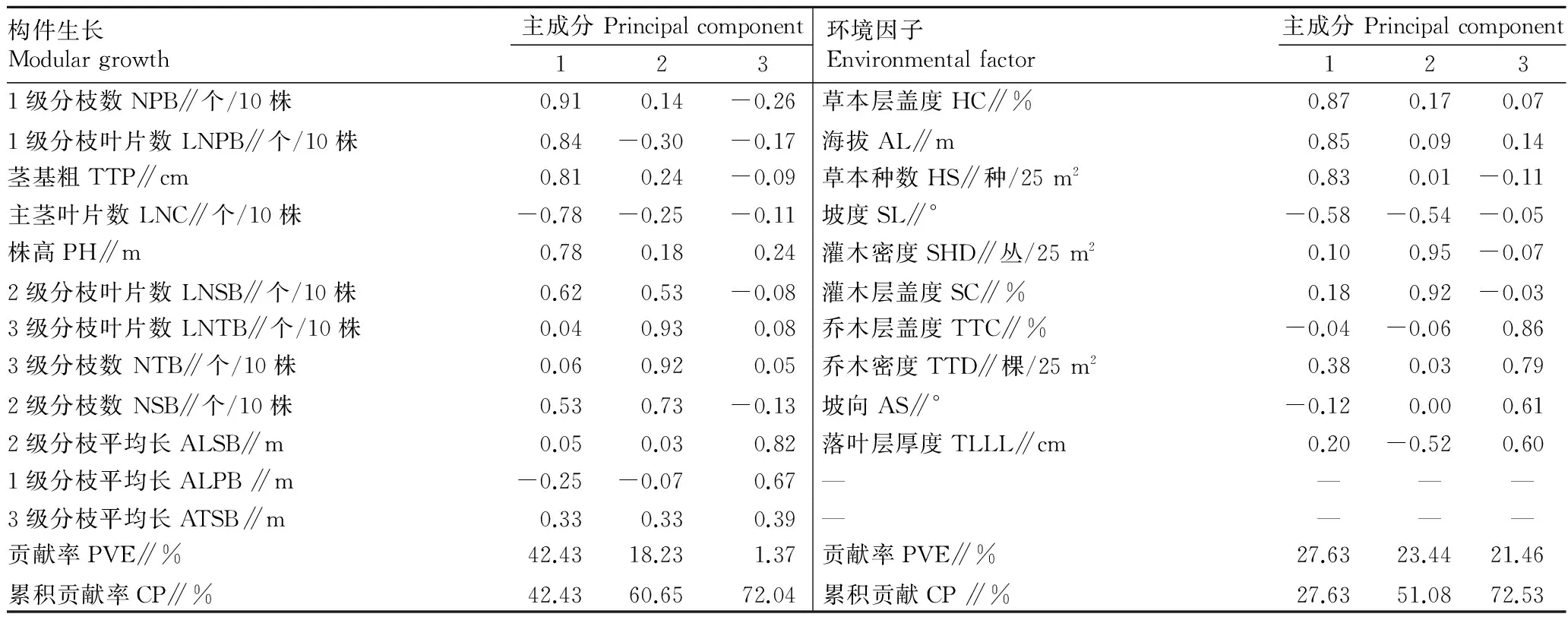
表4 不同生境紫茎泽兰构件生长和环境因子的主成分分析结果
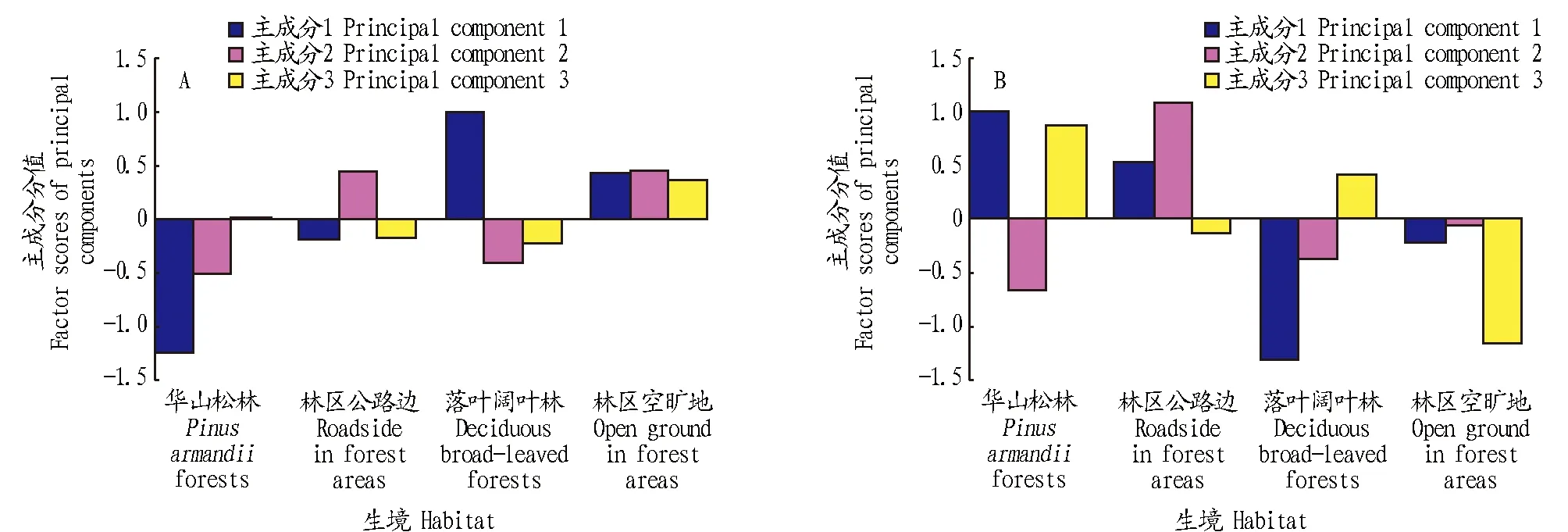
图2 不同生境紫茎泽兰构件生长 (A) 和环境因子 (B) 的主成分分值比较Fig.2 Comparison of the factor scores of growth state (A) and the scores of environmental factors (B) in different habitats of Eupatorium adenophorum
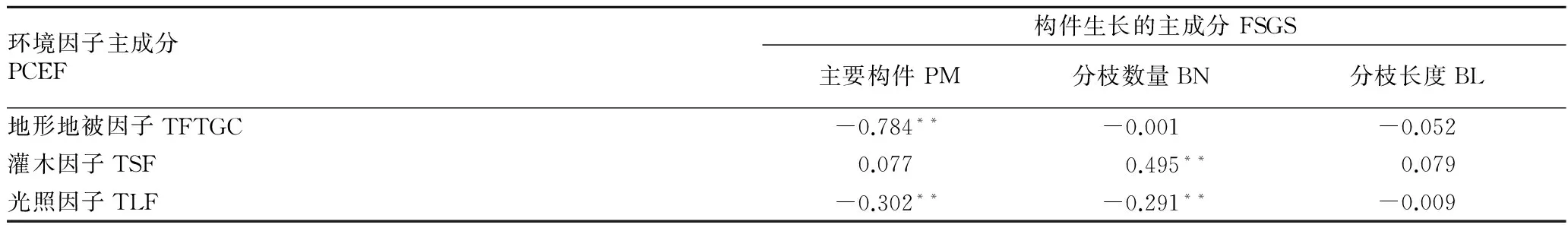
环境因子主成分PCEF构件生长的主成分FSGS主要构件PM分枝数量BN分枝长度BL地形地被因子TFTGC-0.784**-0.001-0.052灌木因子TSF0.0770.495**0.079光照因子TLF-0.302**-0.291**-0.009
注:**表示相关性达到极显著水平(P<0.01)。
Note:** stands for correlation reached extremely significant level(P<0.01).
3.2行为响应的影响因子环境的异质性是影响大中山紫茎泽兰构件生长的关键。该研究中的主成分分析揭示了紫茎泽兰构件生长和环境因子在不同生境中的响应和变化规律。相关性分析表明,生境“地形地被因子”和“光照因子”对紫茎泽兰构件生长有一定的限制性影响。
该研究中华山松林中“地形地被因子”和“光照因子”较大,具体的因子表现为海拔较高、草本层盖度和种数较多,乔木层盖度较大,具有高海拔的气候特征。冬季松林林下,紫茎泽兰幼小植株有冻枯死现象,其他草本的竞争和抑制作用对紫茎泽兰的构件生长也有影响。与其他紫茎泽兰入侵生境相比,松林林冠较郁闭,落叶层厚度大,但凋落的松针难以腐烂,土壤缺乏养分。林下草本虽然较多,但生长相对分散和弱小。松林的温湿和养分条件不利于草本植物的生长,不能充分满足紫茎泽兰生长和分枝所需的光照条件,导致华山松林的紫茎泽兰“主要构件”和“分枝数量”较少。
林区公路边和空旷地的灌木因子较大,光照因子较小,具体的因子表现为灌木密度和盖度大,乔木密度和盖度小。植物主要通过形态变化适应光环境[9]。这2种生境较好的光照条件是造成构件生长的前3个主成分分值较大的主要原因。
落叶阔叶林为旱冬瓜(Alnusnepalensis)林,土壤为黑壤,肥力高,乔木层盖度具季节性变化,林下光照条件也会发生相应改变。该生境“地形地被因子”较小,光照因子较大,表现为海拔较低,草本种数和草本盖度小,乔木盖度大,反映了夏季该生境的低海拔气候特征和弱光照条件。紫茎泽兰是阳性偏阴性植物,种子的萌发和幼苗的生长要求比较稳定的高温高湿条件[2]。因此,林内较好的气候和养分条件是紫茎泽兰“主要构件”较大的原因。光照条件的季节变化,特别是夏季的弱光照条件是该生境“分枝数量”较少的原因。
3.3防治对策利用构件生长的指示作用确定重点防控区。紫茎泽兰在不同环境中的适应响应及由此引起的形状变化,对保护区确定重点防控区及采取相应的对策具有指示和提示作用。例如,在紫茎泽兰长势好的林区空旷地、林区公路边和落叶阔叶林中应加强机械或化学防除,遏制紫茎泽兰进一步蔓延疯长,以防对保护区内的珍稀动植物造成不利影响。
避免或减少对保护区原生生境的人为干扰。扰动生境,如林区空旷地和林区公路边等,坡度较缓,往往受人为干扰较大,这些生境紫茎泽兰大量入侵,生长茂密;相反,在植被保护较好的常绿阔叶林中,紫茎泽兰很少生长,甚至不生长。因此,应尽量保护好区内的原生植被,尤其是常绿阔叶林,避免人为砍伐或破坏,从而达到长期有效的抑制紫茎泽兰入侵、生长和扩张的目的。如果一旦发现原生植被遭轻度破坏,应尽早尽快采取措施恢复。
确定限制性因子,为生态防治对策的形成提供理论依据。该研究表明地形地被因子和光照因子对紫茎泽兰构件生长有一定的限制性影响,如草本较多,乔木盖度较大的生境中紫茎泽兰生长不良。因此,对已出现紫茎泽兰危害的地块,保护区可以通过植被改造、增加群落的郁闭度和种植与紫茎泽兰有竞争力的土著草本,努力营造能有效防治紫茎泽兰入侵和生长的生态防护林。
参考文献
[1] 赵国晶,马云萍.云南省紫茎泽兰的分布与危害的调查研究[J].杂草学报,1989,3(2):37-40.
[2] 刘伦辉,刘文耀,郑征,等.紫茎泽兰个体生物及生态学特性研究[J].生态学报,1989,9(1):66-70.
[3] 卢志军,马克平.地形因素对外来入侵种紫茎泽兰的影响[J].植物生态学报,2004,28(6):761-767.
[4] HUBER H,HUTCHINGS M J.Differential response to shading in orthotropic and plagiotropic shoots of the clonal herbGlechomahirsute[J].Oecologia,1997,112(4):485-491.
[5] 王裕康.南华大中山省级自然保护区综合科学考察报告[R].南华县林业局,2000.
[6] 孙书存,陈灵芝.不同生境辽东栎的构型差异[J].生态学报,1999,29(3):358-364.
[7] FOWLER J,COHEN L,JARVIS P.Practical statistic for field biology[M].West Sussex:Open University Press,1998.
[8] 赵相建,王孝安.太白红杉分枝格局的可塑性研究[J].西北植物学报,2005,25(1):113-117.
[9] NIINEMETS U,TENHUNEN J D.A model separating leaf structural and physiological effects on carbon gain along light gradients for the shade-tolerant speciesAcersaccharum[J].Plant,cell and environment,1997,20(7):845-866.
The Modular Growth ofEupatoriumadenophorumin Different Habitats and Behavioral Responses to Environment
LIU Zhao1,2, LI Wei2, ZHOU Wei2
(1. College of Agriculture and Biotechnology, Hexi University, Zhangye, Gansu 734000; 2. Department of Forestry, Southwest Forestry University, Kunming, Yunnan 650224)
Abstract[Objective] The aim was to explore the response of module growth of Eupatorium adenophorum to changed environments. [Method] One hundred and twenty plots were established respectively in Pinus armandii forest, deciduous broad-leaved forest, roadsides and open ground in forest areas where Eupatorium adenophorum were invaded in Nanhua part of Ailaoshan National Nature Reserve. And data of advantage plants modular of Eupatorium adenophorum and environment factors were measured. The growth status and environmental factors of Eupatorium adenophorum in different habitats were analyzed. Principal component analysis and correlation analysis were adopted to study the relationship between principal component factors of modular growth variables and environmental variables. [Result] The results indicated that there were significant differences (P<0.01) in modular growth of Eupatorium adenophorum advantage plants among different habitats. In Pinus armandii forest, the plants of Eupatorium adenophorum were weaker than other habitats and the allocation in branch class level was unequal, which were closely related to the weak sunlight condition, climatic characters of high elevation and the higher herb coverage. In open ground and roadsides in forest areas, the branching density and intensity of each level branch were higher than other two habitats and the allocation in branch class level were equal, which were related to strong sunlight. In deciduous broad-leaved forests, plants of Eupatorium adenophorum were strong and exuberant and the allocation in branch class level was unequal, which were related to the fertile soil and the weak sunlight condition under canopy in summer. [Conclusion] Environmental change is the key influencing modular growth of Eupatorium adenophorum and modular growth of Eupatorium adenophorum responses different sensitivity to environmental changes. We hereby suggest that the different habitats in the nature reserve should be managed by different manners in order to prevent the invasion of Eupatorium adenophorum and/or decrease its harm when it is invaded.
Key wordsHabitat; Eupatorium adenophorum; Modular growth; Environmental factor; Ailaoshan National Nature Reserve
基金项目国家重点基础研究发展计划项目(2002CB111407);云南省软科学重大项目(2004RK07)。
作者简介刘钊(1981- ),男,山东菏泽人,副教授,博士,从事野生动植物保护与利用研究。
收稿日期2016-03-11
中图分类号S 432.2+2
文献标识码A
文章编号0517-6611(2016)12-020-05
鸣谢野外工作得到了哀牢山国家自然保护区南华分区管理局王学荣副局长和查国富主任、云南省楚雄州自然保护区管理局张仁功先生大力支持,在此一并致谢!
猜你喜欢
落叶果树(2021年6期)2021-02-12
体育科技(2016年2期)2016-02-28
河北渔业(2015年10期)2015-10-15
西华师范大学学报(自然科学版)(2015年3期)2015-02-27
应用海洋学学报(2014年2期)2014-11-26
河北大学学报(自然科学版)(2012年3期)2012-03-25
环保科技(2012年2期)2012-01-10
