
水稻恶苗病菌对咪鲜胺的敏感性及生物学特性的研究
2016-08-07 13:14徐瑶李鹏刘洪亮穆娟微
黑龙江八一农垦大学学报 2016年3期
徐瑶,李鹏,刘洪亮,穆娟微
(1.黑龙江八一农垦大学农学院,大庆163319;2.黑龙江省农垦科学院植物保护研究所)
水稻恶苗病菌对咪鲜胺的敏感性及生物学特性的研究
徐瑶1,李鹏2,刘洪亮2,穆娟微2
(1.黑龙江八一农垦大学农学院,大庆163319;2.黑龙江省农垦科学院植物保护研究所)
为了明确黑龙江省水稻恶苗病菌对咪鲜胺的产生抗性情况,利用菌丝生长速率法检测了来自黑龙江省8个县(市)的32个恶苗病菌株对咪鲜胺的敏感性,确定敏感基线值为0.068 9 μg·mL-1,有15.63%供试菌株为低抗菌株,3.12%为中抗菌株,没有检测到高抗菌株,表明黑龙江省水稻恶苗病菌对咪鲜胺的敏感性明显下降,已经产生抗性菌株。比较了抗性菌株JMS-3和敏感菌株NH-1的生物学特性,发现抗性菌株在生物学特性方面发生了一定变化,抗性菌株JMS-3菌丝生长速度明显低于敏感菌株NH-1,敏感菌株NH-1最适pH为7,抗性菌株JMS-3最适pH为6,敏感菌株NH-1最适氮源为硝酸钠,抗性菌株JMS-3最适氮源为硝酸钾。
水稻恶苗病菌;咪鲜胺;抗药性;生物学特性
水稻恶苗病是由串珠镰孢菌(Fusarium moniliforme Sheld)引起的真菌病害[1],是严重影响黑龙江省水稻生产的主要病害之一,发病地块一般减产10%~20%,严重的可减产50%以上。近年来,随着水稻种植面积的不断扩大、旱育秧技术的推广及种植户在药剂使用技术上存在不足,使得水稻恶苗病的发生日趋严重[2],有关该病的研究得到人们日益重视。
自20世纪90年代咪鲜胺代替多菌灵防治恶苗病以来,咪鲜胺以防效高、对作物安全等优势倍受农民青睐。咪鲜胺浸种防治水稻恶苗病在黑龙江已应用长达20年,连续使用单一杀菌剂会导致抗药性的出现[3],检测其抗药性对保护水稻安全生产意义重大。通过测定黑龙江省不同地区恶苗病菌对咪鲜胺的敏感性,确立该地区恶苗病菌对咪鲜胺的敏感性基线,进而评价该地区恶苗病菌对咪鲜胺的抗性水平,并探讨具有抗药性菌株的生物学特性的变化,为黑龙江省监测恶苗病菌对咪鲜胺的抗药性奠定基础,为指导水稻恶苗病的防控提供理论依据。
1 材料与方法
1.1 供试菌株
于2013年8月在黑龙江省虎林、阿城、讷河、佳木斯、铁力、富裕、绥化和庆安等8个县(市)采集87个水稻恶苗病病株标样,采用组织分离方法进行病原菌的分离,通过单孢分离纯化培养获得62个恶苗病菌菌株,每个县(市)随机抽取4个菌株,共计32个供试菌株。
1.2 恶苗病菌对咪鲜胺敏感性测定
通过预备试验确定出药剂的5个有效浓度,配制含药平板。以室内生长速率法测定咪鲜胺对水稻恶苗病菌菌株的毒力大小[4-5]。在活化后的恶苗病菌菌落边缘上打取直径为5 mm的菌碟,挑取菌碟置于含药平板培养皿的中央,将其置于25℃恒温培养箱中培养,7 d后以十字交叉法测菌落直径,计算不同浓度药剂对各菌株菌丝生长抑制率。求出药剂对菌株的毒力公式,即回归方程y=ax+b及x与y之间的相关系数r,计算出咪鲜胺对各供试菌株的抑制中浓度(EC50)。根据病菌对咪鲜胺的敏感性频率分布建立恶苗病菌对咪鲜胺的敏感基线,确定抗性指数和抗性菌株频率。抗性指数=供试菌株EC50/敏感基线EC50。抗性指数(0,30)为敏感菌株,抗性指数(3,10)为低抗菌株,抗性指数(10,100)为中抗菌株,抗性指数(100,∝)为高抗菌株[6-7]。
1.3 温度对病原菌菌丝生长的影响
设10、15、20、25、30、35、40℃共7个处理,每处理重复3次。将对咪鲜胺敏感菌株和抗性菌株在PDA培养基平板上培养5 d后,在菌落边缘用打孔器取直径7 mm的菌片,然后将菌片接到PDA平板中央,每皿1片,分别置于不同处理温度条件下培养,5 d后测量菌落直径。
1.4 光照对病原菌菌丝生长的影响
设全天24 h黑暗、12 h光照/12h黑暗、全天24 h光照等3个处理,每处理重复3次。将对咪鲜胺敏感菌株和抗性菌株在PDA培养基平板上培养5 d后,在菌落边缘用打孔器取直径7 mm的菌片,然后将菌片接到PDA平板上,每皿1片,接入平板中央位置,分别置于25℃恒温箱中不同光照条件下培养,5 d后测量菌落直径。
1.5 pH对病原菌菌丝生长的影响
用1 mol·L-1HCl和1 mol·L-1NaOH将PDA培养基调成pH值为4、5、6、7、8、9、10、11等8个处理。将各处理分别制成平板,每处理3皿,移对咪鲜胺敏感菌株和抗性菌株直径7 mm菌片于平板中央,置于25℃恒温箱中培养,5 d后测量菌落生长直径。
1.6 不同碳源对病原菌菌丝生长的影响
供试碳源为淀粉、葡萄糖、蔗糖、麦芽糖、果糖。以查氏培养基为基础培养基,以等碳量分别用各供试碳源取代基础培养基内的蔗糖,配置成不同碳源的供试培养基,将不同碳源的培养基倒入灭菌培养皿中制成平板,在平板中央移入在PDA培养基平板上培养直径5 mm的菌片,每皿1片,每处理重复3次,置于25℃恒温箱中培养,7 d后测量菌落直径。
1.7 不同氮源对病原菌菌丝生长的影响
供试氮源为硝酸钠、硫酸铵、硝酸钾、氯化铵、脯氨酸、甘氨酸。以查氏培养基为基础培养基,以等氮量分别用各供试氮源取代基础培养基内的硝酸钠,配置成不同氮源的供试培养基,将不同氮源的培养基倒入灭菌培养皿中制成平板,在平板中央移入在PDA培养基平板上培养直径5 mm的菌片,每皿1片,每处理重复3次,置于25℃恒温箱中培养,7 d后测量菌落直径。
2 结果与分析
2.1 供试菌株对咪鲜胺的敏感性
利用室内生长速率法测定了咪鲜胺对32个供试菌株的毒力大小,通过分析得出各个菌株的毒力回归方程和EC50值,相关系数均在0.917 7以上。各菌株的EC50值差异较大,最小EC50值为0.001 5 μg·mL-1,最大EC50值为1.306 7 μg·mL-1,相差871倍(表1)。
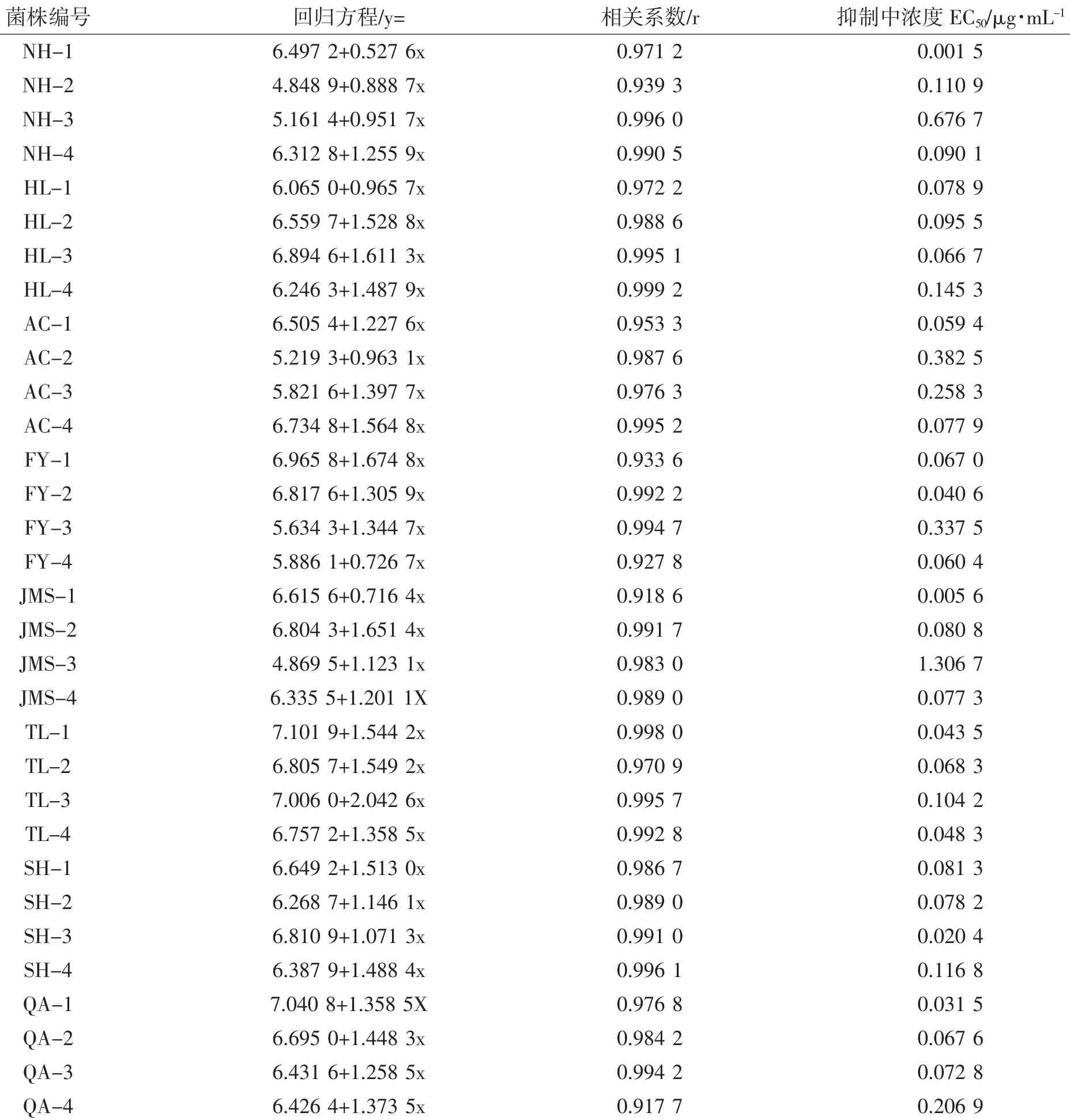
表1 恶苗病菌对咪鲜胺的敏感性Table 1 Sensitivity of Fusarium fujikuroi to prochloraz
依据表1中供试的32个恶苗病菌菌株的EC50值,从0 μg·mL-1开始,以0.05 μg·mL-1为截距,将EC50等分为27个区间,统计每一个区间出现供试菌株频率(表2)。由表2可知,EC50值在0~0.15 μg·mL-1范围内,水稻恶苗病菌对咪鲜胺敏感性的频率分布呈连续正态分布,为敏感菌株群体,其EC50平均值为0.068 9 μg·mL-1,由于无法找到从未用过咪鲜胺的恶苗病菌群体,故实验就以较为敏感菌株群体的EC50平均值(0.068 9 μg·mL-1)作为敏感基线值。依据该敏感基线值计算出各供试菌株的抗性指数并判断其敏感性,在32个供试菌株中,有26个菌株的抗性指数(0,3)为敏感菌株,占81.25%;有5个菌株的抗性指数(3,10)为低抗菌株,占15.63%;有1个菌株抗性指数(10,1000)为中抗菌株,占3.12%;没有检测到高抗菌株。表明黑龙江省水稻恶苗病菌对咪鲜胺的敏感性明显下降,已经产生抗性菌株,在药剂防治方面应及时采取新的防控措施。

表2 恶苗病菌对咪鲜胺敏感性的分布情况Table 2 Distribution of sensitive Fusarium fujikuroi to prochloraz
2.2 抗性菌株在生物学特性方面发生的变化
2.2.1 温度对病菌菌丝生长的影响
温度对抗性菌株JMS-3和敏感菌株NH-1生长具有明显影响,抗性菌株JMS-3和敏感菌株NH-1的菌丝生长温度范围和最适温度是一致的,在10~35℃均能生长,40℃均不能生长,最适温度均为35℃,但在相同温度下抗性菌株JMS-3菌丝生长量明显低于敏感菌株NH-1,在适温30℃条件下培养5 d后,菌落净生长直径分别为49.5和58.9 mm(图1)。
2.2.2 光照对病菌菌丝生长的影响
实验结果表明,病菌在不同光照条件下生长差异不明显,有无光照情况下均能生长,12 h光照/12 h黑暗条件下生长最好。在同等光照条件下,抗性菌株JMS-3菌落净生长直径明显低于敏感菌株NH-1(表3)。
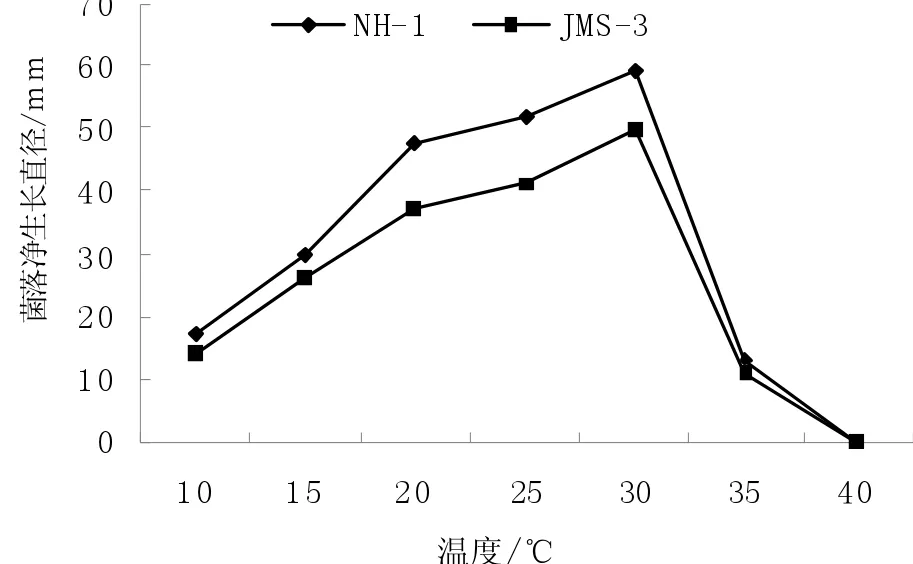
图1 温度对病原菌菌丝生长的影响Fig.1 Effects of temperature to mycelial growth

表3 光照对病原菌菌丝生长的影响Table 3 Effects of light to mycelial growth
2.2.3 pH对病菌菌丝生长的影响
实验结果表明,在pH值4~11范围内病菌均能较好地生长,在相同pH值条件下,抗性菌株JMS-3菌丝生长速度明显低于敏感菌株NH-1。抗性菌株JMS-3在pH 6时菌丝生长最快,培养5 d后,菌落净生长直径40.1 mm;而敏感菌株NH-1在pH 7时菌丝生长最快,培养5 d后,菌落净生长直径49.7 mm(图2)。
2.2.4 不同碳源对病菌菌丝生长的影响
从表4可以看出,病菌在淀粉为碳源的培养基上生长最快,蔗糖次之。在相同碳源培养基上,抗性菌株JMS-3菌丝生长量明显低于敏感菌株NH-1(表4)。

图2 pH对病原菌菌丝生长的影响Fig.2 Effects of pH on mycelial growth
2.2.5 不同氮源对病菌菌丝生长的影响
实验结果表明,抗性菌株JMS-3和敏感菌株NH-1对氮源需求有较大差别,抗性菌株JMS-3菌丝在以硝酸钾为氮源的培养基上生长最快,培养7 d后菌落净生长直径为50.8 mm,脯氨酸次之。敏感菌株NH-1菌丝在以硝酸钠为氮源的培养基上生长最快,培养7 d后菌落净生长直径为76.8 mm,硝酸钾次之。在相同氮源培养基上,抗性菌株JMS-3生长速度明显低于敏感菌株NH-1(表5)。
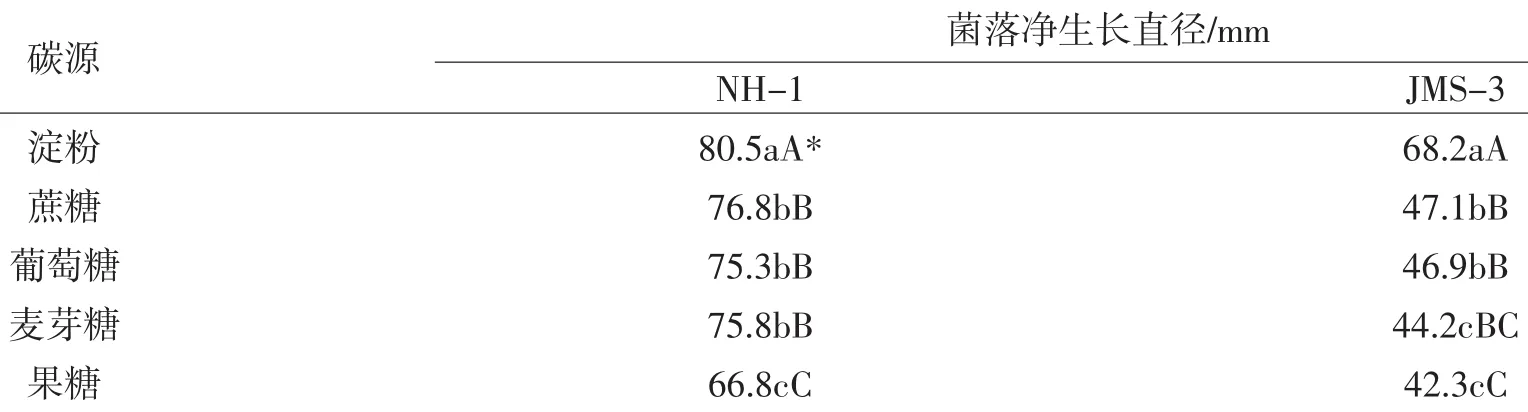
表4 不同碳源对病原菌菌丝生长的影响Table 4 Effects of carbon sources to mycelial growth

表5 不同氮源对病原菌菌丝生长的影响Table 5 Effects of nitrogen sources to mycelial growth
3 结论与讨论
赵志华等[8]研究表明,水稻恶苗病菌对咪鲜胺具有潜在的抗性风险;刘永锋等[9]研究表明,田间恶苗病菌对咪鲜胺已有一定的抗性;杨红福等[10]报道由于长期单一使用咪鲜胺浸种防治水稻恶苗病,导致水稻恶苗病菌对咪鲜胺抗药性大幅度上升,抗性菌株成为田间优势种群,并导致田间咪鲜胺对水稻恶苗病丧失防效。通过检测来自黑龙江省8个县(市)的32个恶苗病菌株对咪鲜胺的敏感性,发现有81.25%菌株表现敏感,18.75%菌株已经产生抗药性,抗性菌株的产生是咪鲜胺浸种的防治效果降低的主要原因。继续使用咪鲜胺浸种防治水稻恶苗病存在很大的风险,建议更换作用机制不同的药剂推广使用,以控制恶苗病的发生与危害。
对抗性菌株JMS-3和敏感菌株NH-1比较研究发现,抗性菌株生物学特性方面发生了一定变化。与敏感菌株NH-1相比,抗性菌株JMS-3菌丝生长速度明显降低,生长条件更喜偏酸(pH值为6),最适氮源为KNO3。综合分析认为水稻恶苗菌菌丝在10~35℃均能生长,最适生长温度为30℃;最适菌丝生长的pH呈弱酸性或中性;光暗交替条件有利于菌丝生长;菌丝适于以淀粉、硝态氮为碳氮源的培养环境下生长。产祝龙等[11]研究表明,人工诱导抗咪鲜胺突变菌株的菌丝生长速率小于敏感菌株;Tasleen-uz等[12]研究指出抗咪鲜胺菌株的生长速率比敏感菌株要低,这与研究结果相同。水稻生产上大量施用氮肥、土壤pH适宜,恶苗病发病率较高,而适当使用有机肥,改善土壤环境,可减少发病率,温度对恶苗病菌丝生长具有明显影响,避免中午高温时插秧,以减轻发病,春季气温较高时应加强对恶苗病的防治,避免恶苗病的大发生。
[1]王拱辰.水稻恶苗病病原菌的研究[J].植物病理学报,1990,20(2):93-98.
[2]产祝龙,丁克坚,檀根甲.水稻恶苗病的研究进展[J].安徽农业科学,2002,30(6):880-883.
[3]杨谦.植物病原菌抗药性分子生物学[M].2版.北京:科学出版社,2011.
[4]张亚玲,王宝玉,台莲梅,等.黑龙江省和吉林省稻瘟病菌种群多样性研究[J].黑龙江八一农垦大学学报,2014,26(1):1-4.
[5]陈夕军,卢国新,童蕴慧,等.水稻恶苗病菌对三种浸种剂的抗性及抗药菌株的竞争力[J].植物保护学报,2007,34(4):425-430.
[6]兰波,李湘民.江西省稻瘟病菌对富士一号的抗药性研究[J].江西农业大学学报,2007,29(3):351-355.
[7]袁杰,杨学辉.贵州省稻瘟病菌对三环唑敏感性研究[J].西南农业学报,2006,19(2):219-221.
[8]赵志华,张锡明,范洁茹,等.咪鲜胺对水稻恶苗病菌及其抗药突变体生长发育的影响[J].农药学学报,2007,9(3):251-256.
[9]高宇.水曲柳主要病虫害及其防治方法[J].长春大学学报,2014,24(10):1384-1388.
[10]杨红福,吉沐祥,姚克兵,等.水稻恶苗病菌对咪鲜胺产生的抗药性研究及治理[J].江西学报,2013,25(6):94-96.
[11]产祝龙.水稻恶苗病的防治机理及其病原菌抗药性的研究[D].合肥:安徽农业大学,2003.
[12]Khan T Z,G iliM A,Khan M G.Inheritance studies for bakanae disease of rice[J].Pakistan Journal of phytopathology,1999,11(2):181-182.
Research on Sensitivity and Biological Characteristics of Fusarium fujikuroi to Prochloraz
Xu Yao1,Li Peng2,Liu Hongliang2,M u Juanwei2
(College of Agronomy,Heilongjiang Bayi Agricultural University,Daqing 163319;2.Institute of Plant Protection,Heilongjiang Academy of Land Reclamation Sciences)
In order to clarify Fusarium fujikuroi resistant to prochloraz,the mycelium growth rate method was used to test sensitivity of 32 Fusarium fujikuroi isolates to prochloraz that collected from 8 countries(cities)in Heilongjiang province,and the sensitive baseline value was 0.068 9 μg·mL-1.The low level resistant strains and the medium level resistant strains accounted for 15.63%and 3.12%,respectively,and the high level resistant strains was not detected.The results showed that the sensitivity of Fusarium fujikuroi to prochloraz in Heilongjiang province was significantly lower and resistant strains was produced.Compared the biological characteristics of resistant strains JMS-3 and sensitive strains NH-1,the some changes had taken place in the biological characteristics of resistant strains,and resistant strains JMS-3 mycelium growth rate was significantly lower than sensitive strains NH-1.The optimal pH of sensitive strains NH-1 was 7 and the optimal pH of resistant strains JMS-3 was 6.The optimal nitrogen sources for sensitive strains NH-1 was NaNO3and the optimal nitrogen sources for resistant strains JMS-3 was KNO3.
Fusarium fujikuroi;prochloraz;fungicide resistance;biological characteristics
S482.2+99
A
1002-2090(2016)03-0012-05
10.3969/j.issn.1002-2090.2016.03.003
2015-03-25
黑龙江垦区一戎水稻科技奖励基金会支持项目(水稻包衣剂防治恶苗病配套技术研究);黑龙江垦区一戎水稻科技奖励基金会支持项目(水稻恶苗病菌对咪鲜胺抗药性风险评价及防治新技术示范)。
徐瑶(1988-),女,黑龙江八一农垦大学农学院2012级硕士研究生。
穆娟微,女,研究员,硕士研究生导师,E-mail:mujuanwei@126.com。
猜你喜欢
世界农药(2022年8期)2022-09-02
陕西农业科学(2022年2期)2022-03-28
浙江农业学报(2021年5期)2021-05-31
农药科学与管理(2019年7期)2019-11-29
江西农业大学学报(2018年6期)2019-01-14
新农业(2017年15期)2017-09-11
上海农业学报(2017年4期)2017-04-10
中国酿造(2016年12期)2016-03-01
中国酿造(2014年9期)2014-03-11
植物营养与肥料学报(2011年6期)2011-10-24
