
干旱胁迫对日本落叶松生理指标的影响1)
2016-08-08 00:56王骞春陆爱君冯健于世河陈明文胡艳
东北林业大学学报 2016年8期
王骞春 陆爱君 冯健 于世河 陈明文 胡艳
(辽宁省林业科学研究院,沈阳,110032) (国有抚顺县温道林场) (抚顺县林业局)
干旱胁迫对日本落叶松生理指标的影响1)
王骞春陆爱君冯健于世河陈明文胡艳
(辽宁省林业科学研究院,沈阳,110032)(国有抚顺县温道林场)(抚顺县林业局)
摘要利用盆栽和持续干旱处理及复水后对日本落叶松生长的影响,研究日本落叶松针叶的水饱和亏(WS,D)、相对电导率(E)、丙二醛(MDA)质量摩尔浓度、叶绿素质量分数、超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)酶活性、脯氨酸质量分数和可溶性蛋白质量浓度等抗旱相关生理指标的变化规律。结果表明:随着干旱胁迫程度的增加,水饱和亏、相对电导率、丙二醛质量摩尔浓度、脯氨酸质量分数和可溶性蛋白质量浓度显著上升,其中水饱和亏和相对电导率呈梯度上升趋势,可溶性蛋白质量浓度第6天达到峰值,丙二醛质量摩尔浓度和脯氨酸质量分数在第9天时达到峰值。超氧化物歧化酶、过氧化物酶活性呈上升趋势,超氧化物歧化酶活性第12天达峰值,过氧化物酶活性第9天达峰值。叶绿素a和叶绿素(a+b)质量分数呈先上升后下降趋势,都在第9天达到峰值,第12天出现下降,低于对照。叶绿素b质量分数呈微弱的下降趋势,w(叶绿素a)∶w(叶绿素b)呈上升趋势,第9天达峰值。叶绿素(a+b)质量分数变化主要是由叶绿素a质量分数变化引起的。过氧化氢酶酶活性呈先上升后下降趋势,第6天达峰值,9 d后开始下降且低于对照。用主成分分析法从指标中共提取了两个因子,累计贡献率为91.267%,因子1反映了日本落叶松针叶保水能力、抗氧化酶活性、质膜稳定性和渗透调节物对抗旱性起支配作用,因子2反映叶绿素质量分数对抗旱性起支配作用。通过灰色关联度分析,日本落叶松存活率与各指标变化幅度的关联度从大到小次序为w(叶绿素a)∶w(叶绿素b)、超氧化物歧化酶、丙二醛质量摩尔浓度、过氧化物酶、水饱和亏、脯氨酸、相对电导率、叶绿素b、叶绿素a、叶绿素(a+b)、过氧化氢酶。综合以上结果,把水饱和亏、电导率、脯氨酸和丙二醛质量摩尔浓度作为日本落叶松抗旱的有效指标。
关键词干旱胁迫;日本落叶松;生理指标
干旱胁迫作为一种最普遍的树木逆境形式,影响树木的生长发育,严重时可降低林木的产量和品质[1]。由于植物的物种多样性和基因多样性,树木在长期进化演变中逐渐形成了适合自身生理和生态特征的耐旱机制[2]。树木受到干旱胁迫时,生理指标会发生变化,反映树木受伤害的程度以及耐旱调节能力[3-4]。前人对植物抗旱指标做过大量的研究和报道,如水分生理指标(水分饱和亏、叶水势、相对含水量等),代谢指标(净光合速率、蒸腾速率、呼吸速率等),酶和渗透调节物(SOD、POD、CAT、脯氨酸、可溶性糖和可溶蛋白等),质膜过氧化物丙二醛以及叶绿素质量分数,都可作为鉴定植物抗旱性的重要指标[5]。
研究树木干旱胁迫下的生理指标的变化规律,有助于抗旱机理的研究和耐旱品种的选育。日本落叶松(Larix kaempferi (Sieb.etZucc.)Gord.)具有适应性强、早期速生、成林快、病虫害少、材质优良的特点,是南方亚高山地区和北方地区造林的重要树种[6]。北方地区春季容易发生春旱,春季造林时如果降雨量偏少,会严重影响落叶松造林成活率。本文对不同程度的干旱胁迫对日本落叶松水饱和亏、膜透性、丙二醛、SOD、POD、CAT、脯氨酸、叶绿素质量分数和可溶蛋白质量浓度等生理指标进行研究,揭示日本落叶松在干旱胁迫下的变化规律及对干旱胁迫的适应机制和耐受性。应用主成分分析和灰色关联度分析方法,研究了生理指标与抗旱性的关系,为落叶松抗旱性鉴定提供参考。
1材料与方法
试验材料为清原县大孤家国家级林木良种基地培育的2年生日本落叶松实生苗,于2014年5月,选取生长良好一致的单株移栽入白色塑料花盆中,花盆高30cm,直径20cm,土壤为腐殖质土与珍珠岩按相同比例混合,每盆土壤体积和质量一致,进行常规水分管理,土壤体积含水量控制在20%左右。
1.1干旱胁迫处理
7月初将盆苗移入辽宁省林业科学研究院塑料大棚内,水分管理按常规水分管理要求浇水。于7月21日将36盆生长一致的盆栽苗进行干旱胁迫,设置4个水分处理(断水0、6、9和12d),以断水0为对照(CK)。每个处理设3次重复,3株为1重复,胁迫结束后取各处理苗木针叶进行各项指标测定,每个重复测每株数据1次,取平均值。试验期间沈阳多为晴或多云天气,日照充足,平均每日最低温度为20 ℃,最高温度30 ℃,空气湿度50%~65%。
1.2测定指标及方法
水饱和亏:采取饱和称质量法[7],取同一部位40枚针叶,称鲜叶质量,然后清水浸泡24h后,擦干表面水分,称饱和鲜叶质量。烘箱105 ℃杀青30min,然后置于烘箱80 ℃烘干10h,称干质量。
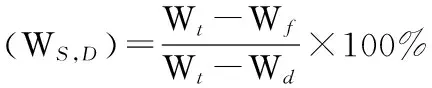
式中:Wt为叶饱和后鲜质量;Wf为叶鲜质量;Wd为叶干质量。
相对电导率:采用电导仪法[7],取10mL蒸馏水于试管中,用DDSJ-308A电导率仪测量其电导率(E0);取0.2g针叶清水冲洗一遍后,用去离子水冲洗3次,吸干表面水分,剪碎、混匀放入试管蒸馏水中,封住试管口,置于室温环境下浸泡24h,测电导率(E1);然后将试管沸水浴30min,冷却至室温后再测浸提液电导率(E2)。
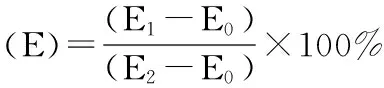
叶绿素质量分数测定:采用分光光度法[8],避光称取剪碎、混合后的新鲜针叶0.1g,放入研钵中,加少量石英砂及3mL体积分数为95%的乙醇,研磨成匀浆,再加乙醇10mL,继续研磨至组织变白,避光静置3~5min。将滤液过滤到25mL棕色容量瓶中,冲洗研钵、玻璃棒数次并用乙醇定容至25mL,摇勻。将叶绿素倒入比色皿中,于665、449mn下比色。按下列公式计算叶绿素a,叶绿素b和叶绿素(a+b)质量分数。
叶绿素a质量分数=(13.7D665-5.76D649)×V/(1 000×W),
叶绿素b质量分数=(25.8D649-7.6D665)×V/(1 000×W),
叶绿素(a+b)质量分数=(6.1D665+20.04D649)×V/(1 000×W)。
式中:D665、D649分别为相应波长下的光密度值;V为提取液的体积;W为叶片鲜质量。
复水后存活率:采样结束后立即复水,按常规水分管理7d后调查苗木存活情况;落叶松苗木复水7d后针叶依然凋萎、发脆枯死脱落,茎杆干缩无韧性被视为已死亡。
复水后存活率=存活苗数量/总苗数量×100%。
1.3其他生理指标测定
游离脯氨酸(Pro)含量采用酸性茚三酮法测定[9];过氧化物酶(POD)活性采用愈创木酚法,超氧化物歧化酶(SOD)活性采用氮蓝四唑光还原法,过氧化氢酶(CAT)活性采用可见光分光光度计法[9];丙二醛(MDA)质量摩尔浓度采用硫代巴比妥酸法[8];可溶蛋白含量用考马斯亮蓝法[9]。
1.4数据处理
数据采用MicrosoftExcel2007和SPSS21.0统计分析软件包进行处理。以单因素方差分析(onewayANOVA)和最小显著差异法(LSD)分析不同胁迫程度下生理指标的差异。
2结果与分析
2.1干旱胁迫对落叶松生理生化指标的影响
2.1.1复水后存活率
从表1可以看出,日本落叶松盆栽苗CK和6d处理复水后存活率为100%,断水9d后复水存活率为67%,断水12d后复水存活率下降为56%。结果表明,断水胁迫6d,日本落叶松可通过自身的调节,表现出一定的抗旱性,避免受到胁迫伤害,但是断水9d后胁迫程度超出了日本落叶松的自我调节范围,造成一定程度的不可逆转的伤害。
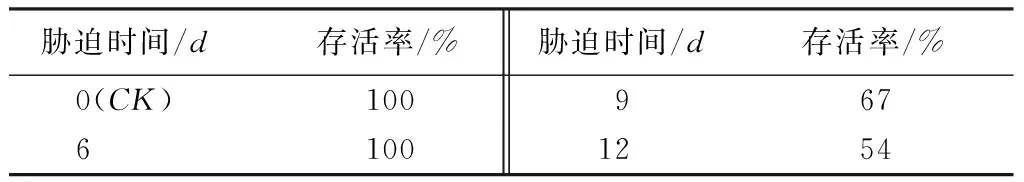
表1 不同干旱胁迫时间复水后成活率
2.1.2不同胁迫时间对水饱和亏、相对电导率、丙二醛的影响
水饱和亏数值越大,表示植物组织水分亏缺越严重,可反映植物叶片的保水复水能力[10]。本试验结果显示(表2),随着干旱胁迫程度的增加,日本落叶松的水饱和亏呈明显的梯度上升趋势。断水6、9和12d的水饱和亏比CK分别上升了64%、221%和346%。经过LSD(α=0.05)多重比较,各处理间水饱和亏差异均达到显性水平。可以看出,落叶松针叶在不同程度的干旱胁迫期间,均出现明显失水现象,说明日本落叶松针叶保水能力较弱。
干旱胁迫下,植物电导率的的变化,可以反映质膜的受害程度。干旱胁迫引起叶片内自由基积累,导致质膜的过氧化,细泡膜结构受到破坏,膜透性增大导致细胞内电解质外渗,使细胞相对电导率增大[11]。由表2可知,日本落叶松针叶相对电导率,随着胁迫程度的增加呈逐渐上升趋势。相对CK,断水6、9和12d的电导率上升了18%、39%和90%。经多重比较,CK和6d的处理与断水12d的处理差异显著。试验结果说明,不同程度的干旱胁迫都对日本落叶松质膜系统造成了伤害,胁迫程度低于9d时,电导率上升幅度相对较小,质膜受伤害程度增加不明显,但胁迫9d后,电导率迅速增加,说明断水9d后膜系统结构开始收到严重破坏。
丙二醛是质膜过氧化后的产物,干旱胁迫后丙二醛的质量摩尔浓度可以反映细胞受损伤的程度[12]。由表2可以看出,随着干旱胁迫程度增加日本落叶松的丙二醛质量摩尔浓度总体呈上升趋势,各胁迫处理的丙二醛质量摩尔浓度分别比CK上升了90%、126%和106%,经多重比较,CK与各断水处理均差异显著,断水处理间无显著差异。干旱胁迫开始6d时,丙二醛质量摩尔浓度就迅速增加,并且与对照呈显著性差异,说明此阶段日本落叶松质膜质膜明显发生了过氧化,胁迫第9天丙二醛质量摩尔浓度达最大值,说明此时质膜过氧化程度达到最大。
2.1.3不同胁迫时间对SOD、POD、CAT活性的影响
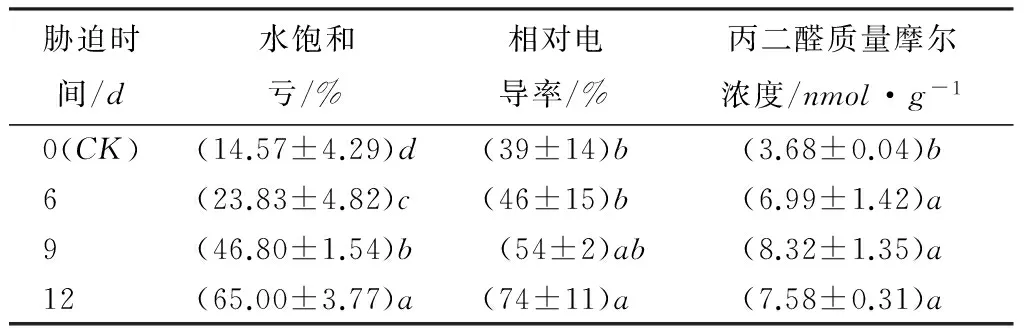
表2 干旱胁迫对水饱和亏、相对电导率、丙二醛的影响
注:表中数据为平均值±标准差;同列不同小写字母表明不同胁迫时间同一指标差异显著(p<0.05)。
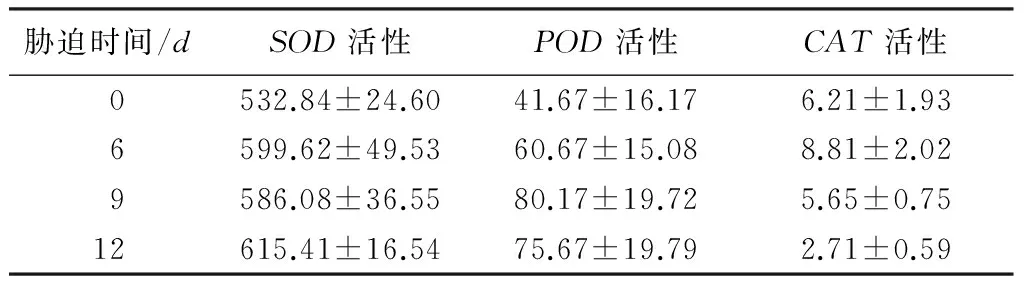
表3 干旱胁迫对SOD、POD、CAT活性的影响U·g-1
注:表中数据为平均值±标准差。
2.1.4不同胁迫时间对叶绿素质量分数的影响
一般研究认为干旱胁迫可以影响叶片色素含量[15-16〗。如表4所示,本试验中,不同处理的日本落叶松叶绿素a和叶绿素(a+b)质量分数,随着胁迫程度的增加呈先上升后下降趋势,但变化幅度较低。与CK相比,胁迫6和9d,叶绿素a质量分数增加了3%和3.4%,胁迫12d时降低了0.7%。叶绿素(a+b)质量分数断水6和9d时比CK增加了1.7%和2%,断水12d比CK减少了2%,叶绿素(a+b)质量分数变化主要由叶绿素a变化引起。叶绿素b质量分数呈梯度降低趋势,各阶段的叶绿素b质量分数分别降低了1.7%、1.75%和5.28%。w(叶绿素a)∶w(叶绿素b)值呈先上升后降低趋势,各胁迫时期比CK各增加了4.8%、5.3%和4.9%。叶绿素a质量分数、叶绿素(a+b)质量分数和w(叶绿素a)∶w(叶绿素b)值各处理间均无显著差异。结果说明不同程度干旱胁迫对日本落叶松叶绿素质量分数产生了影响,胁迫9d内叶绿素质量分数变化不明显,只在胁迫12d时出现了相对较大的下降幅度。
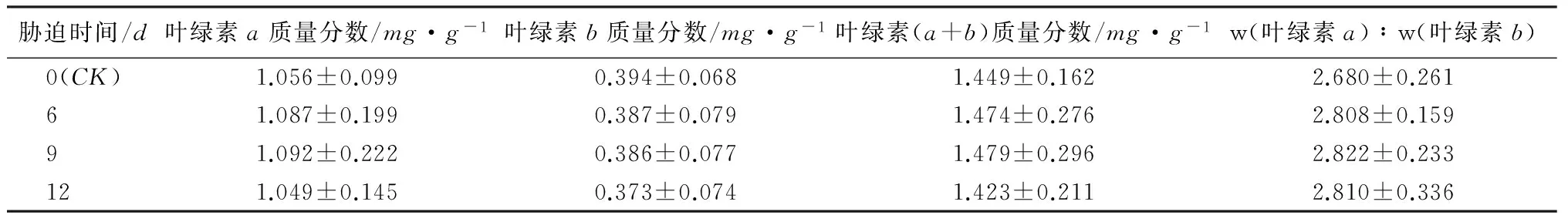
表4 干旱胁迫对叶绿素质量分数的影响
注:表中数据为平均值±标准差。
2.1.5不同胁迫时间对脯氨酸、可溶性蛋白的影响
在植物中,渗透调节物质在发生干旱胁迫时,可以降低细胞渗透势和水势,保护膜系统,维持细胞代谢正常[17]。如表5所示,在本试验中,受到干旱胁迫期间,日本落叶松组织的脯氨酸含量呈先上升后下降的趋势,在断水9d时达最大值,分别是CK的3倍、9倍和8倍,其中断水9和12d时与CK差异显著。可溶蛋白在受胁迫期间表现为上升—下降—上升的变化趋势,在断水6d时达最大值。与CK相比,分别上升了55%、27%和36%,其中断水6d与CK差异显著。可以看出,发生干旱胁迫6d时,可溶蛋白质量浓度显著上升,脯氨酸含量也有所增加,但变化不显著。胁迫9d后可溶蛋白质量浓度开始下降,脯氨酸质量分数显著增加。
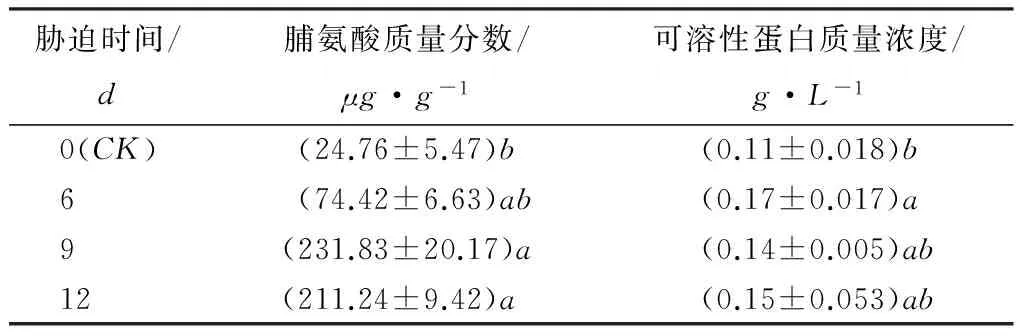
表5 干旱胁迫对脯氨酸、可溶性蛋白的影响
注:表中数据为平均值±标准差;同列不同小写字母表明不同胁迫时间同一指标差异显著(p<0.05)。
2.2主成分分析
为了全面评价各指标对日本落叶松抗旱性的贡献程度,运用主成分分析法提取主要因子。共提取了2个因子,累计贡献率为91.267%,说明提取的因子包含的信息可以反映各生理指标原始特征参数的大部分信息,具有代表性(表6)。对因子旋转后得到因子载荷矩阵。因子1中载荷绝对值较大的指标,从大到小依次为水饱和亏、过氧化物酶、电导率、超氧化物歧化酶、叶绿素b、脯氨酸、丙二醛和w(叶绿素a)∶w(叶绿素b),分别为0.950、0.932、0.923、0.911、0.901、0.898、0.892和0.871,反映了日本落叶松针叶保水能力、抗氧化酶活性、质膜稳定性和渗透调节物对抗旱性起支配作用。因子2中荷绝对值较大的指标,从大到小依次为叶绿素a、叶绿素(a+b)和过氧化氢酶,分别为0.976、0.926和0.761,反映叶绿素质量分数对抗旱性起支配作用。而可溶性蛋白在这两个因子中的载荷都较小,只有0.594和0.555。
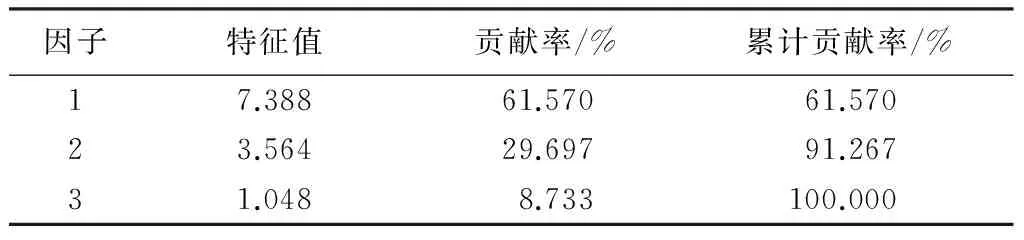
表6 指标因子分析结果
2.3灰色关联度分析
为研究日本落叶松干旱胁迫过程中各生理指标的的变化幅度与植株受害程度的关系,以断水时间为时段,将主成分分析中提取的载荷大于0.6的指标相对于CK的增减幅度作比较数列,将断水后复水成活率作为参考数列,组成灰色系统,按分辨系数为0.5分析各生理指标变化幅度与成活率的关联度。各指标变化幅度与成活率的关联程度由大到小依次为w(叶绿素a)∶w(叶绿素b)、超氧化物歧化酶、丙二醛、过氧化物酶、水饱和亏、脯氨酸、相对电导率、叶绿素b、叶绿素a、叶绿素(a+b)、过氧化氢酶,关联系数分别为0.940 2、0.926 7、0.919 9、0.886 2、0.846 4、0.845 2、0.842 7、0.841 5、0.834 4、0.562 5和0.541 7。说明植株受害程度与w(叶绿素a)∶w(叶绿素b)值变化幅度关联度最密切,其次依次是超氧化物歧化酶、丙二醛、过氧化物酶等指标。
3结论与讨论
植物的水饱和亏是反映干旱条件下叶片持水情况的生理指标,可以衡量植物的耐旱性[18]。
魏良民[19]通过比较梭梭和白榆等植物发现,同样生长环境和时间条件下,中生植物的水饱和亏明显大于旱生植物,抗旱能力较强的梭梭的水饱和亏,在水分充足的5月到干旱的7、8月的变化幅度也最低,因此可认为抗旱性强的树种,干旱胁迫时水饱和亏低于抗旱性弱的树种,而且随着干旱程度的增加,变化幅度也比较小。本研究中,日本落叶松水饱和亏随胁迫时间的增加,表现出显著增加趋势,表明日本落叶松叶片失水较快,保水能力较弱。尽管不同种类的落叶松的叶片构造有差异[20],但研究发现,兴安落叶松与长白落叶松等落叶松属植物的叶片都存在保水能力差,缺乏抗脱水的能力,属于低水势忍耐脱水耐旱树种[21-22],日本落叶松也应属于此类树种。
研究表明,树木受干旱胁迫时膜透性增加,电导率上升,并且干旱胁迫后抗旱性强的树种电导率增加幅度低于抗旱性相对较弱的树种[4,23]。本研究中,日本落叶松电导率随胁迫强度的增加也呈现逐渐上升的趋势,增加幅度较高,说明日本落叶松在干旱胁迫下维持膜稳定性能力较弱。
抗旱性不同的树种或同一树种的不同种源,不同种类的抗氧化酶活性变化幅度规律并不相同。邱真静等[24]发现,不同种源沙拐枣在干旱胁迫下,超氧化物歧化酶活性增加幅度由大到小为武威、张掖、酒泉,过氧化物酶增加幅度由大到小为酒泉、武威、张掖。申学圣等[25]对岷江上游干旱河谷区的不同树种的研究中发现,干旱条件下,超氧化物歧化酶的活性增幅由大到小为刺槐、榆树、油松、岷江柏,过氧化物酶活性增幅为榆树、刺槐、油松、岷江柏。本研究中日本落叶松在不同阶段干旱胁迫下,过氧化物酶和过氧化氢酶活性在收受到胁迫初期变化幅度较大,较为敏感,超氧化物歧化酶活性变化平稳。过氧化氢酶活性6d时达最大值,6d后显著降低,低于CK,说明过氧化氢酶活性较早被抑制,后期主要是超氧化物歧化酶和过氧化物酶发挥抗氧化作用。超氧化物歧化酶活性与CK比最高增幅为15%,呈现双峰曲线,最低值出现在胁迫9d时,过氧化物酶活性与CK比最高增幅为92%,呈现单峰曲线,最高值出现在胁迫9d时,说明在干旱胁迫下日本落叶松抗氧化保护酶是互补和协调共同发挥作用的[26]。
干旱胁迫过程中,抗旱性强的品种比抗旱性弱的品种丙二醛质量摩尔浓度上升的幅度低[27-28],这种情况可能与植物清除丙二醛的能力有关[29]。本研究中,日本落叶松的丙二醛质量摩尔浓度在第6天时与CK出现显著差异,增幅达到90%,说明干旱胁迫前期日本落叶松清除丙二醛的能力较弱。
受到干旱胁迫时,植物体内产生的超氧自由基对叶绿素产生破坏,引起植物叶绿素质量分数降低[30]。有研究发现干旱胁迫前后叶绿素变化幅度小的树种抗旱性相对较强[31]。本研究中,胁迫6和9d时日本落叶松叶绿素a和叶绿素(a+b)质量分数高于CK,分析原因可能是由于叶片失水导致的浓缩现象,在胁迫12d时日本落叶松叶绿素a和叶绿素(a+b)质量分数低于CK,说明叶绿素受破坏程度加剧,含量降低现象已经不受浓缩现象影响。叶绿素b质量分数在各个阶段呈梯度下降趋势,说明日本落叶松受干旱胁迫时叶绿素b对超氧自由基反应比叶绿素a敏感。
植物在干旱胁迫下受到超阴离子活性氧的伤害,引起膜系统过氧化,光合系统和代谢紊乱最终引起植体受到伤害甚至死亡,因此,植物受伤害程度和存活率可以反映植物的抗旱性。本研究中,通过日本落叶松干旱胁迫过程中不同时期的存活率与各指标的增减幅度的灰色关联度分析发现,与日本落叶松的w(叶绿素a)∶w(叶绿素b)值变化幅度与抗旱性关系最密切,其他指标变化幅度与抗旱性关联程度依次为超氧化物歧化酶、丙二醛、过氧化物酶、水饱和亏、脯氨酸和相对电导率。通过对日本落叶松抗旱相关指标的分析,利用主成分分析法发现,日本落叶松针叶叶绿素含量、保水能力、抗氧化酶活性、质膜稳定性和渗透调节物对抗旱性起支配作用。
综上所述,受到干旱胁迫时日本落叶松抗脱水能力、维持膜的稳定性能力和清除丙二醛能力都较弱,受胁迫短期内可以通过渗透调节和抗氧化酶保护作用维持正常生长,但随着干旱胁迫加剧,因为光合作用受阻,质膜受破坏严重,水份大量流失,体内酶活受到抑制导致生长受抑制甚至死亡。经过综合分析将水饱和亏、电导率、脯氨酸质量分数和丙二醛质量摩尔浓度可以作为评价日本落叶松抗旱性的主要指标。
参考文献
[1]LANGEOL,KAPPENL,SCHULZEED.Waterandplantlife:problemsandmodernapproaches[M].Berlin:HeidelbergsSpringerverlag,1976.
[2]李吉跃.植物耐旱性及其机理[J].北京林业大学学报,1991,13(3):92-100.
[3]谢寅峰,沈惠娟,罗爱珍,等.南方7个造林树种幼苗抗旱生理指标的比较[J].南京林业大学学报,1999,23(4):13-16.
[4]张卫华,张方秋,张守攻,等.3种相思幼苗的抗旱性研究[J].林业科学研究,2005,18(6):695-700.
[5]刘萍,康峰峰,王旭航.林木抗旱鉴定指标及数量分析方法研究进展[J].河南林业科技,2006,26(2):20-23.
[6]马常耕.落叶松种和种源选择[M].北京:农业大学出版社,1992.
[7]王韶唐,荆家海,丁钟荣,等.植物生理学实验指导[M].西安:陕西科学技术出版社,1986.
[8]郝再彬,苍晶,徐仲.植物生理实验[M].哈尔滨:哈尔滨工业大学出版社,2004.
[9]熊庆娥,植物生理实验教材[M].成都:四川科学技术出版社,2003.
[10]韩艳,林夏珍.5种常绿阔叶树幼苗的抗旱性比较[J].浙江林学院学报,2009,26(6):822-828.
[11]贾根良,代惠萍,冯佰利,等.PEG模拟干旱胁迫对糜子幼苗生理特性的影响[J].西北植物学报,2008,28(10):2073-2079.
[12]许桂芳.PEG胁迫对2种过路黄抗性生理生化指标的影响[J].草业学报,2008,17(1):66-70.
[13]蒋明义,郭绍川.水分亏缺诱导的氧化胁迫和植物的抗氧化作用[J].植物生理学通讯,1996,32(2):144-150.
[14]胡义,胡庭兴,陈洪,等.干旱胁迫及复水对香樟幼树生理特性及生长的影响[J].西北植物学报,2015,35(2):0294-0301.
[15]夏阳.水分逆境对果树脯氨酸和叶绿素含量变化的影响[J].甘肃农业大学学报,1998,28(1):26-31.
[16]黄承建,赵思毅,王龙昌,等.干旱胁迫对苎麻叶绿素含量的影响[J].中国麻业科学,2012,32(5):208-212.
[17]郭华军.水分胁迫过程中的渗透调节物质及其研究进展[J].安徽农业科学,2010,38(15):7750-7753,7760.
[18]浦锦春,刘家琼,刘新民,等.我国荒漠植物的水分饱和亏[J].中国沙漠,1989,9(3):44-53.
[19]魏良民.几种旱生植物水分生理特性的比较研究[J].新疆农业大学学报,1991,8(2):75-79.
[20]王翠华,刘慧昌,张颂云,等.落叶松不同种和日本落叶松不同家系叶片横切面构造及性状的研究[J].辽宁林业科技,1993(6):1-5.
[21]张建国,宋廷茂,李志丹,等.兴安落叶松和樟子松苗木水分关系的研究(Ⅲ)[J].北京林业大学学报,1993,15(增刊1):38-46.
[22]冯玉龙,王文章,敖红.长白落叶松和樟子松等5种树种抗旱性的比较[J].东北农业大学学报,1998,26(6):16-19.
[23]谢安德,唐春红,潘启龙,等.干旱胁迫对不同种源麻风树幼苗生理特性的影响[J].江西农业大学学报,2011,33(2):0306-0311.
[24]邱真静,李毅,种培芳,等.基于PEG胁迫响应的不同地理种源沙拐枣抗旱性评价[J].中国沙漠,2011,30(5):1231-1237.
[25]申学圣,宫渊波,陈林武,等.岷江上游干旱河谷4个乡土树种的抗旱生理指标主成分分析[J].贵州林业科技,2012,40(4):1-6.
[26]祈娟,徐柱,王海青,等.旱作条件下披碱草属叶的生理生化特征分析[J].草业学报,2009,18(1):39-45.
[27]陈少裕.膜脂过氧化与植物逆境胁迫[J].植物学通报,1989,6(4):211-217.
[28]吴涛,陈少瑜,彭明俊,等.不同种源膏桐在干旱胁迫下生理指标的变化[J].西北林学院学报,2008,23(2):7-11.
[29]于金慧,柏明娥,方伟,等.干旱胁迫对4种灌木生理生化特性的影响[J].浙江林学院学报,2009,26(4):485-489.
[30]蒋明义,杨文英,徐江,等.渗透胁迫下水稻幼苗中叶绿素降解的活性氧损伤作用[J].植物学报,1994,36(4):289-295.
[31]潘昕,邱权,李吉跃,等.干旱胁迫对两种速生树种叶绿素含量的影响[J].桉树科技,2013,30(3):17-22.
第一作者简介:王骞春,男,1979年7月生,辽宁省林业科学研究院,高级工程师。E-mail:3140373@qq.com。
收稿日期:2015年12月31日。
分类号Q945.78
EffectsofDroughtStressonPhysiologicalIndicesofJapaneseLarch//
WangQianchun,LuAijun,FengJian,YuShihe
(LiaoningAcademyofForestrySciences,Shenyang110032,P.R.China);ChenMingwen(TheStateForestFarmofFushunCounty);HuYan(ForestryBureauofFushunCounty)//JournalofNortheastForestryUniversity,2016,44(8):13-17,40.
Apotexperimentwasconductedtoinvestigatetheeffectsofcontinuouslydroughtstressonphysiologicalindexesincludingwatersaturationdeficit(WSD),relativeconductivity,malondialdehyde(MDA),Chlorophyll,superoxidedismutaseSOD,peroxidasePOD),catalase(CAT),proline(Pro)andsolubleprotein(SP)ofJapaneselarch.Withtheincreaseofthedegreeofdroughtstress,WSD,RC,MDAcontent,ProcontentandSPcontentwereincreasedsignificantly,WSDandRCwasgradientupwardtrend,thecontentofSPreachedthepeakonthe6thday,thecontentsofMDAandProreacheditspeakonthe9thday.TheactivitiesofSODandPODwasincreasedwitharisingtrend,andthatofSODreachedthepeakonthe12thdayandthe9thday,respectively.Thecontentsofchlorophyllaandchlorophyll(a+b)increasedfirstlyandbothreachedthepeakonthe9thday,anddecreasedonthe12thday,lowerthanthatofCK.Thecontentsofchlorophyllbshowedaslightdecreasetrend,andchlorophyll(a/b)showedarisingtrend,andreachedapeakonthe9thday.Thecontentsofchlorophyll(a+b)weremainlycausedbythechangeofchlorophyllacontent.TheactivityofCATincreasedfirstlyreachedthepeakonthe6thday,anddecreasedtoCKafter9d.Fromthephysiologicalindexeswithprincipalcomponentanalysismethod,twofactorswereextractedwiththecumulativecontributionrateof91.267%.Factor1reflectedtheJapaneseLarchneedlesholdingwaterability,anti-oxidaseactivityandstabilityofplasmamembraneandosmoticadjustmentagainstdroughtcontrolfunction,andFactor2reflectedtheleafchlorophyllcontentagainstdroughtcontrolfunction.ThegreycorrelativedegreeofsurvivalratewithotherindexeswasinthedescendingrankofChl(a/b),SOD,MDA,POD,WSD,Pro,RC,Chlb,Chla,Chl(a+b),andCAT.Therefore,WSD,Chlcontent,theactivitiesofSODandPOD,andMDAcanbeusedaseffectiveindicatorsfordroughtresistanceidentification.
KeywordsDrought stress; Larix kaempferi; Physiology indices
1)“十二五”国家科技支撑计划项目(2012BAD01B01)。
责任编辑:潘华。
猜你喜欢
吉林农业·下半月(2017年1期)2017-02-16
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
安徽农学通报(2017年1期)2017-02-15
物联网技术(2016年12期)2017-01-21
安徽农学通报(2016年24期)2017-01-12
山东农业科学(2016年11期)2016-12-17
