
田间麦蚜和实验室麦蚜抗药性毒力测定和生化测定的研究
2016-09-06 09:07李宏德
甘肃农业 2016年2期
李宏德
(甘肃省定西市安定区种子管理站,甘肃 定西 743000)
田间麦蚜和实验室麦蚜抗药性毒力测定和生化测定的研究
李宏德
(甘肃省定西市安定区种子管理站,甘肃定西743000)
麦蚜是危害小麦的主要害虫,主要通过取食和病毒传播来损害农作物。麦蚜的防治一直以化学防治为主,但由于其繁殖快,遗传背景复杂等,致使其抗性发展速度快、程度高,对多种常用农药已产生了显著抗性。笔者对从田间带回的麦蚜进行室内培养,并从其生测,生物化学角度对其抗性机理进行了研究,其结果如下:㈠分别用四大类别7种农药对田间麦蚜和室内麦蚜进行了毒力测定,在所有的药剂中,氧化乐果对室内品系的致死中浓度最低,毒死蜱对室内品系的致死中浓度最高。相对于室内品系,田间麦蚜对氧化乐果产生了40倍的抗性,对菊酯类的敌杀死产生了11倍的抗性,对灭多威产生了3倍的抗性,对于辛硫磷,吡虫啉,速灭杀丁,毒死蜱等抗性不明显。㈡测定了田间麦蚜和室内品系的乙酰胆碱酯酶和羧酸酯酶的酶活性和动力学常数。田间麦蚜的乙酰胆碱酯酶的活性及Vmax均小于敏感品系,而Km值大于敏感品系。对于其羧酸酯酶的水解活性,当以α-NA为底物时,田间麦蚜的羧酸酯酶水解活性明显低于室内品系的,而动力学常数Km值与Vmax均大于室内品系;当以β-NA为底物时,田间麦蚜的羧酸酯酶水解活性,动力学常数Km,Vmax与室内麦蚜无明显区别。
麦蚜;羧酸酯酶;乙酰胆碱酯酶;抗性机制;玻璃管药膜法
一、概述
麦蚜是危害小麦的主要害虫,麦蚜俗称小麦腻虫,麦蚜是甘肃省麦田发生的主要虫害之一,主要发生种类有麦二叉蚜(SchizaphisgraminumRonbani)、麦长管蚜 (Macrosiphumavenae Fabricius)二种,其次还有麦无网长管蚜和禾缢管蚜等发生危害较轻的种类。麦蚜1年可发生10~20余代。麦蚜在适宜的环境条件下,都以无翅型孤雌胎生若蚜生活。在营养不足、环境恶化或虫群密度大时,则产生有翅型迁飞扩散,但仍行孤雌胎生,只是在寒冷地区秋季才产生有性雌雄蚜交尾产卵。卵来春孵化为干母,继续产生无翅型或有翅型蚜虫。卵长卵形,长为宽的1倍,约1 mm,刚产出的卵淡黄色,逐渐加深,5d左右即呈黑色。干母、无翅雌蚜和有翅胎生蚜外部形态亦相似,除具性器外,一般个体稍小。麦蚜在寄主茎、叶及嫩穗上刺吸为害,吸取汁液使叶片出现黄斑或全部枯黄,生长停滞,分蘖减少,籽粒饥瘦或不能结实,对产量影响较大。麦二叉蚜还可传播小麦病毒病。有些地方麦蚜发生高峰从2次增加到3次,防治次数多,用药量加大。目前对麦蚜的防治仍以化学防治为主,化学农药无节制的滥用,造成该虫的抗药性迅速增加。所以对麦蚜抗药性的研究就变的尤为重要。
(一)麦蚜的抗药性现状
魏岑等在1983~1987采用6个省市共8个地区麦蚜种群对6种农药进行了点滴法毒力测定,探明了甘肃张掖、高台麦长管蚜对氧化乐果产生了10~15倍的抗性,随后其他省区也陆续报道了麦蚜对氧化乐果的抗性,且产生抗性的种类也不断增多,包括常用的有机磷和氨基甲酸酯类药剂,其中特别对氰戊菊酯和溴氰菊酯的抗性最为突出。
苏建昆等1986~1989年采用点滴法测定了扬州市郊区麦蚜对常用杀虫剂的敏感度水平,禾谷缢管蚜对各种杀虫剂的敏感程度依次为溴氰菊酯>久效磷>乐果>氧化乐果>对硫磷>马拉硫磷>氰戊菊酯>敌百虫。麦长管蚜对各种杀虫剂的敏感程度依次为久效磷>马拉硫磷>氧化乐果>对硫磷>氰戊菊酯>敌百虫。2种麦蚜对各种杀虫剂的敏感度有一定的同步关系。但禾谷缢管蚜对久效磷和氧化乐果的敏感度分别下降10和7倍。
刘慧平等1991~1995年采用点滴法测定了山西主要麦区晋中、临汾、运城麦长管蚜对拟除虫菊酯杀虫剂溴氰菊酯、氰戊菊酯、氯氰菊酯,甲氰菊酯和有机磷杀虫剂氧化乐果、乐果、马拉硫磷、辛硫磷、敌敌畏的敏感性。测定结果表明:晋中麦长管蚜已对氰戊菊酯和氯氰菊酯产生了一定水平的抗性,抗性指数分别为8.4和9.4倍。而对有机磷杀虫剂除临汾、晋中地区对氧化乐果和乐果产生了低水平抗药性,抗性指数为5~7倍。
潘文亮等于2000年进行了吡虫啉对麦长管蚜的抗药性研究,结果发现,所测不同地区的麦长管蚜对吡虫啉的敏感性差异不大,主要原因是由于各地尚未大量使用吡虫啉防治这两种害虫。
洪波等于2001~2002年采用点滴法测定了宁夏地区麦长管蚜、禾谷缢管蚜、麦二叉蚜对几种杀虫剂的敏感性。结果表明,3种麦蚜对灭多威、吡虫啉、丁硫克百威敏感性高,而对敌敌畏、氯氰菊酯、氧化乐果和烟碱敏感性较低。比较不同地区麦蚜种群对杀虫剂的敏感性,各地区麦长管蚜种群对丁硫克百威、灭多威的敏感度差异较大,对其它供试药剂的敏感度差异相对较小。
王晓军等2004年采用麦穗浸渍法在室内测定了麦长管蚜和禾谷缢管蚜对吡虫啉可湿性粉剂和乳油的敏感度。结果表明,禾谷缢管蚜对吡虫啉的敏感度是麦长管蚜的3~4倍(基于LC50的比较)。在北京、河南、江苏、湖北、四川等地同浓度的吡虫啉对禾谷缢管蚜的防治效果要好于麦长管蚜,特别是在低浓度时尤为明显[1-5]。
(二)抗药性机理的研究
害虫抗药性是杀虫剂选择的结果,抗性机理可分为行为抗性、生理抗性和生化抗性(代谢抗性)3种,行为抗主要有2类:依赖刺激的行为回避和非依赖刺激的行为回避,生理、生化特性机理主要有以下几个方面:表皮穿透速率敏感度降低,靶标部位敏感度降低或改变,代谢酶活性升高。关于麦蚜的抗性机理问题,一般认为,药物在渗入表皮后达到作用部位靶体(如神经系统)之前,药物的新陈代谢增加变成无毒的结果。这种新陈代谢包含了一系列的酶,其中水解酶是主要的一种。有机磷农药是磷酸酯类化合物,具有酯基,可能被水解酶断裂而失去毒力,水解酶的含量和活性越高,分解断裂有机磷的能力越强,农药毒力越低。吡虫啉是内吸性杀虫剂,同时具有胃毒和触杀作用,该药对飞虱、粉虱、蚜虫等刺吸式口器害虫及其抗药性种群,具有优异的防治效果,害虫抗吡虫啉的机理目前尚未明确。从现有结果来看,抗性的形成可能与害虫解毒能力提高和靶标部位对毒素敏感性降低有关,或可能伴随着表皮穿透率降低。Choi等研究了桃蚜对吡虫啉的抗性机理,利用14C-吡虫啉研究了抗性和敏感品系体内的残留动态,与敏感品系相比,抗性品系体内的吡虫啉残留量较少,对吡虫啉的排泄量有所增加。抗吡虫啉桃蚜品系的乙酰胆碱酯酶活性是敏感种群的1.4倍,但吡虫啉对2个品系都没有抑制作用PBO和异稻瘟净增效试验表明,吡虫啉和PBO按1∶1和1∶5混合处理抗性个体,增效比值分别是69.4和250。吡虫啉和IBP按 1∶1和1∶5混合处理抗性个体,增效比值分别是227和80.6,而对敏感品系的增效比值都在0.6~1之间。以α-NA和β-NA为底物测定了酯酶活性,抗性品系分别是敏感品系的1.4和2.4倍。以上研究表明,P450单加氧化酶和酯酶解毒能力的提高与抗性形成有关。
(三)羧酸酯酶与有机磷抗药性分子机制
1.羧酸酯酶概述。羧酸酯酶(Carboxlesterase,CaEs,EC3.1.1.1)属于α/β折叠水解酶家族,在水存在的条件下能够水解代谢带羧基的酯。α/β折叠水解酶包括磷酸三酯酶(如对氧磷酶)、胆碱酯酶、脂肪酶以及许多酶。这些酶的命名是基于它们8对几乎平行的α/β结构及其具有典型的可塑性的特性。蛋白质的这种折叠属性使其结构域大小的变化范围很大。羧酸酯酶存在于许多组织中,包括肝,肾,小肠,心脏,肌肉,肺,脑,睾丸,鼻和呼吸组织,脂肪组织,血细胞和血液。然而,羧酸酯酶的表达和活性依赖于组织,不同组织表达水平和活性有很大的变化。[6-10]
昆虫中羧酸酯酶在细胞中可能主要以可溶性蛋白分子形式存在,羧酸酯酶分布的研究较少,在黑尾叶蝉、棉蚜、白背飞虱和褐飞虱中羧酸酯酶活性主要分布于上清液中(姚洪渭等,2001)。在铜绿蝇敏感品系和马拉硫磷抗性品系中,羧酸酯酶活力均主要集中于上清液和线粒体中(Whyardetal,1994)。李腾武等(1999)对采自不同地区的小菜蛾种群幼虫不同亚细胞层羧酸酯酶的活性分布结果表明,羧酸酯酶活性亚细胞分布没有明显差异,羧酸酯酶活性以线粒体层分布最高,占34%~52%。
CaEs可催化水解众多药物和内源化合物,如盐酸肾上腺素异戊酯、碳酸盐、可卡因、水杨酸、辣椒素、棕榈酸酰、氟哌啶醇、咪唑普利、吡咯烷生物碱、类固醇等。近年来研究发现CaEs还能水解老鼠胰腺和肾中类固醇中的脂肪酸酯[11-16]。此外,CaEs又被确定为代谢活化前药过程中的一个关键性酶,可起到抑制药物和活化前药的作用[5]。
有关羧酸酯酶多态性已被报道了,其中值得一提的是鉴别了单个羧酸酯酶(CaE2或CES2)中15个单核苷酸多态性(SNPs)。随着众多酯酶被鉴定,酯酶家族一直不断的壮大。Aldridge(1953)根据酯酶与有机磷的相互作用把酯酶分为三类:一是A-酯酶,这类酶能耐受有机磷,并把它们作为底物进行水解;二是B-酯酶,这类酶对对氧磷等有机磷很敏感,能被这些有机磷抑制;三是C-酯酶,既不被有机磷抑制,又不能水解这些有机磷。许多A酯酶能通过活性位点上乙酰化的半胱氨酸水解有机磷,这些酶被称为磷酸三酯水解酶(Reiner,1993;Walker,1993)。羧酸酯酶主要指B-酯酶,这些酶具有一个丝氨酸活性位点,因此B酯酶和丝氨酸水解酶是同义的。
2.羧酸酯酶与有机磷抗药性。Matsumura和Brown在1961年首次报道了抗马拉硫磷跄斑库蚊(Calextarsalls)的羧酸酯酶活性高于敏感品系。世界各地报道的许多昆虫对有机磷的抗性往往与高活性酯酶有关。抗性和敏感个体酯酶活性的极大差异可以是量的差异,也可以是质(酯酶发生变构)的差异,或两者兼有。关于酯酶在有机磷抗性棉蚜中的作用机理,Owusu(1994)认为CarE很可能永久地与杀虫剂结合,使杀虫剂失效进而保护杀虫剂作用的活性位点。Suzuki(1993)认为CarE对杀虫剂的解毒作用主要是酶与杀虫剂的结合作用,CarE的所有同工酶都与有机磷抗性有关,其总酶活力与抗性程度有关。Satio(2000)通过杀螟硫磷对CarE活力的完全抑制这一实验结果印证了Suzuki的观点。
(四)乙酰胆碱酯酶与有机磷抗药性分子机制
1.乙酰胆碱酯酶概述。胆碱酯酶(cholinesterase,ChE)是一类丝氨酸水解酶,同其他丝氨酸水解酶相比,在活性中心丝氨酸周围有一个共同的五肽序列,并在胆碱酯酶和甲状腺球蛋白的羧基端区之间具有明显的序列同源性。依其底物特异性可分为乙酰胆碱酯酶和丁酰胆碱酯酶(BuChE)两种。在脊椎动物体内,胆碱有两种:AChE和BuChE,两者的基因结构、氨基酸序列及空间三维构型均显示了很高的同源性,两者的主要差异在于对乙酰胆碱(ACh)、丁酰胆碱(BuCh)两种底物有明显不同的选择性。昆虫体内,胆碱酯酶仅有一种,该酶对底物的选择性位于AChE和BuChE之间。但对ACh的选择性仍强于BuCh,所以昆虫体内的胆碱酯酶仍习惯性地称为AChE。AChE主要分布于动物的神经系统和脊椎动物的红血球以及肌肉中。AChE的体躯分布因昆虫种类而异。Booth等(1987)分析了八种昆虫包括嶂螂、蟋蟀、蜜蜂、家蝇、蚊子、皮蠢、刺蜡和纳的AChE的组织分布,发现其遍布于中枢神经系统,包括突触和非突触结构。在鞘翅目的棉铃象甲(Anthonomusgrandis)、棉铃虫(Helicoverpa armigera)、二化螟(Chilosuppressalis)、黑腹果蝇(Drosophila melanogast)、骚扰角(Haematohia irritans)中,AChE主要分布在头胸部,以腹部最少(Bulland Lundquist,1968;Melanson等,1985;Xu and Bull,1994;高希武等,1998;彭宇,2000)。对于蜚镰目的美洲大镰和白背飞虱(Sogatella furcifera)其头部、胸部、腹部身体的各部分AChE活性相差不大(唐振华,1993;姚洪渭等,2001)。另外,在一些昆虫的血淋巴(Arpagaus and Toutant,1985)中也发现了AChE的存在。可见,AChE不仅存在于昆虫的中枢神经系统,也存在于其它的组织器官。
2.乙酰胆碱酯酶靶标不敏感与有机磷抗药性。乙酰胆碱酯酶(acetylcholinesterase,AChE)是生物神经传导中的一种关键性的酶。在胆碱能突触间,该酶能降解乙酰胆碱,终止神经递质对后膜的刺激作用,保证神经冲动在突触间正常传导。AChE是有机磷和氨基甲酸酯类杀虫剂的作用靶标。有机磷和氨基甲酸酯类杀虫剂首先和AChE形成一个复合体,然后发生酶的磷酰化或氨基甲酰化,这也就造成了酶的被抑制。AChE被抑制后,使ACh在突触处积累,过量的ACh造成去极化阻断,从而抑制了正常的神经传导,神经传导的阻断必然会影响整个生理生化过程的失调与破坏,最终造成昆虫的死亡。
在抗性昆虫,AChE活性往往增强,而更多是由于AchE变构所致。也有研究认为抗性与AchE蛋白多态性的丢失相关[17-23]。
乙酰胆碱酯酶靶标不敏感性是害虫对有机磷和氨基甲酸酯类杀虫剂产生抗药性的主要原因(Feyereisen1995)。自从首次在黑腹果蝇(Drosophila melanogaster)乙酰胆碱酯酶基因中发现与抗药性有关的突变位点(F115S,I199V,G303A,F368Y)后(Muteroet al.,1994),随后又在马铃薯叶甲(Leptinotarsa decemlineata)(Zhu et al.,1996;Kim et al.,2006),家蝇(Musca domestica)(Walsh et al.,2001),桔小实蝇(Bactrocera dorsalis)(Hsu et al.,2006),棉蚜 (Aphis gossypii)(Li and Han 2004;Andrews et al.,2004;Toda et al.,2004;Benting and Nauen 2004),麦长管蚜(Sitobion avenae)(Chen et al.,2007),桃蚜(Myzus persicae)(Nabeshima et al.,2003),淡色库蚊(Culexpipiens)(Weill et al.,2003;Haoues et al.,2007),疟蚊(Anopheles gambiae)(Weill et al.,2003),三带喙库蚊(Culextritaeniorhynchus)(Oh et al.,2006),二斑叶螨(Tetranychus urticae)(Anazawa et al.,2003),棉铃虫(Helicoverpa armigera)(Ren et al.,2002),苹果蠹蛾(Cydia pomonella)(Stefano et al.,2006),Bractocera oleae(Vontas et al.,2002),小菜蛾(Jietal.,2005;Lee etal.,2007)的Ⅰ型或Ⅱ型乙酰胆碱酯酶中发现一些与抗药性相关的突变位点。
(五)研究意义
目前国内外科学家对麦蚜的生物学、生态学、防治等方面报道较多,而对抗药性的研究仅限于表层的研究,如对农药的敏感性,致死中浓度的研究较多。而对抗药性机理,分子蛋白等方面的报道相对来说较少。所以本实验通过对两种种群麦蚜的毒力测定及羧酸酯酶和乙酰胆碱酯酶活性及动力学参数进行测定对比,明确了羧酸之酶和乙酰胆碱酯酶在麦蚜产生抗性过程中所起的作用。为田间麦蚜的抗性诊断及治理提供可靠的理论依据和切实可行的方法。
二、麦蚜的毒力测定
(一)材料与方法
1.供试药剂。75%氧乐果原药(原中国人民解放军第九七一五工厂),40%辛硫磷乳油 (江苏宝灵化工股份有限公司),40%毒死蜱乳油(江苏宝灵化工股份有限公司),20%灭多威可湿性粉剂(英国LK作物科学有限公司),20%速灭杀丁,25%敌杀死,20%灭多威,95%吡虫啉。
2.田间麦蚜与敏感麦蚜的选育
(1)供试虫源。田间麦蚜是从田间麦田随机采集带有麦蚜叶片,带回室内,并在不接触药剂的情况下进行室内饲养。室内麦蚜为实验室在不接触药剂下长期饲养虫源.
(2)麦蚜的饲养。在上方为玻璃、四周为100目尼龙纱围住的养虫笼中单独饲养田间麦蚜和敏感麦蚜,养虫笼基本可以避免外界昆虫的进入,麦苗正常饲养,所用培养土为营养土。每盆土中种20~30粒小麦种子,于正常室温光照下饲养,待麦苗长到一定高度,将麦蚜均匀的接到麦苗上,继续培养,每天浇水一次,当麦蚜长成无翅成蚜时即可选用。
(3)毒力测定方法。毒力测定采用Shotkoski等(1990)和高希武等[16]的玻璃管药膜法,略有改进。先将杀虫药剂用丙酮配置成1 000 mg/L的母液,使用时用丙酮稀释,按等比或等差稀释成需要的浓度[16]。从稀释好的药液中吸取200 μL加入到直径2 cm,高5.2 cm的玻璃管(内表面积36 cm2)中,将玻璃管放于桌上均匀滚动,待丙酮挥发后用于毒力测定。对照单独用丙酮处理。挑取健康一致的无翅成蚜进行试验,每个制备好的药膜管放置20头蚜虫,将试管口用棉花堵住,防止蚜虫跑出。蚜虫挑选时,应在所有麦苗上均匀选取,不应只在一盆麦苗上挑选,这样不能代表整个种群,实验结果势必不会准确。麦蚜尽量选择个头较大,大小较一致的无翅成蚜。在室内正常饲养条件下(温度18~25℃,相对湿度大约为50%~70%,光照为17L∶7D)[16],3 h后检查死亡率,其中只有一条腿动或者完全不动者记为死,记录死亡数,用polo软件计算出LC50,斜率等。每个药剂六七个浓度,每个浓度3次重复。
本实验中先对7种农药进行预实验,分别记录每种农药对田间麦蚜和敏感麦蚜致死的大概中心浓度.以此浓度为中间值,上下共设六组浓度梯度,彼此间为2倍的关系,稀释时所用溶剂为丙醇,每组设3次重复。
(二)结果与分析
实验结果如下表所示,田间麦蚜和室内麦蚜对新烟碱类药剂吡虫啉的LC50值分别为0.4548和1.63417 mg/L;对氨基甲酸酯类药剂灭多威LC50值为2.19450和0.73026 mg/L;对有机磷类药剂氧乐果,辛硫磷、毒死蜱的LC50值0.586~18.336 mg/L和0.045~9.45 mg/L。田间麦蚜对7种农药的敏感度由高到低依次为吡虫啉,毒死蜱,辛硫磷,灭多威,速灭杀丁,氧化乐果,敌杀死。室内麦蚜对7种农药的敏感度由高到低为氧化乐果,灭多威,吡虫啉,敌杀死,辛硫磷,速灭杀丁,毒死蜱
田间麦蚜对三种药剂的抗性相对室内麦蚜来说非常明显,其中,对于氧化乐果,敌杀死,灭多威三种农药的LC50,田间麦蚜的LC50分别为室内麦蚜的40倍,11倍,3倍。由表1可看出,田间麦蚜对于有机磷类的氧化乐果的抗性最高。

表1 田间麦蚜与敏感麦蚜的毒力测定结果
(三)讨论
关于蚜虫的生物测定方法有浸虫法、浸叶片法、点滴法、喷雾法等多种,本实验为了便于操作、节省时间,选择了玻璃管药膜法。处理后检查死亡率的时间是影响实验结果的一个重要因子,通过观察不同时间的死亡率,最后确定了处理后3 h作为标准。玻璃管药膜法在国外也是一种常用的药剂敏感度测定方法。
点滴法是一种比较精确的方法,但是其致命的缺点是要求测定人员技术熟练,特别是对蚜虫类的测定,在国外很少采用;叶片药膜法由于小麦叶片比较狭窄不适于用于叶片药膜法;浸虫法的程序也是比较复杂的。相比之下玻璃管药膜法易用性要明显优于其他方法[16]。
对两类麦蚜毒力测定结果可看出,田间麦蚜相对于室内品系来说对有机磷类农药的氧化乐果抗性最高,而对同是有机磷类的毒死蜱和辛硫磷致死中浓度很低,非常敏感。对于氨基甲酸酯类的灭多威有一定的抗性。对于菊酯类的敌杀死抗性很高,而对于同是菊酯类的速灭杀丁来说敏感度较高。由此可见,田间麦蚜对于某些杀虫剂有明显抗性,而对于一些杀虫剂来说其敏感度较高。
三、羧酸酯酶和乙酰胆碱酯酶与田间麦蚜抗药性关系研究
羧酸酯酶(CarE)是昆虫体内重要的水解酶之一,一般认为它在有机磷及拟除虫菊酯杀虫剂抗性中起着重要的作用。乙酰胆碱酯酶(AChE)作为有机磷和氨基甲酸酯类药剂的作用靶标,在害虫抗药性形成过程中具有重要作用。了解羧酸酯酶和乙酰胆碱酯酶特性的变化对研究麦蚜的抗性机制有着重要的作用。
(一)材料与方法
1.药剂。75%氧乐果原药(原中国人民解放军第九七一五工厂);α-乙酸萘酯(α-NA)、β-乙酸萘酯(β-NA)、固蓝B盐、毒扁豆碱和牛血清白蛋白(BSA)均购自 SIGMA-ALDRICH,考马斯亮蓝G-250。
2.试虫来源。同上。
(二)乙酰胆碱酯酶比活力测定
1.试剂配制。配置0.1 mg/mL的牛血清蛋白液;磷酸缓冲液(pH7.0,0.04M)。
考马斯亮蓝G-250试液的配置:考马斯亮蓝G-250 100 mg+95%乙醇50 ml+85%H3PO4100 mL,蒸馏水定容至1 L,过滤。
配置ATCH(硫代乙酰胆碱)10 mm/l;DTNB(5,5-二硫双-2-硝基苯甲酸)0.0125 mm/l。
2.乙酰胆碱酯酶酶液的制备。取田间麦蚜和敏感麦蚜各300头,在冰水浴中预冷20 min,于4℃,10 000 g冷冻离心20 min,取上清,离心后,若有脂肪体可抽取过滤,抽滤瓶应预先冷冻。将上清液分装于预冷好的离心管中,备用。
3.蛋白质(pr)标准曲线制作。参照Bradford[22]考马斯亮蓝G-250法将试管分成8组,每组设3个重复,按照表2加入试剂。
Buffer为不含Triton x-100(聚乙二醇辛基苯基醚)的PH7.5磷酸缓冲液。大量的去污剂如Triton X-100等会干扰测定。混匀,室温放置10~20 min,分光光度计595 nm处测定吸光值,重复三次取平均值。以标准蛋白浓度(mL)为横坐标,OD值为纵坐标,做出蛋白质标准曲线。
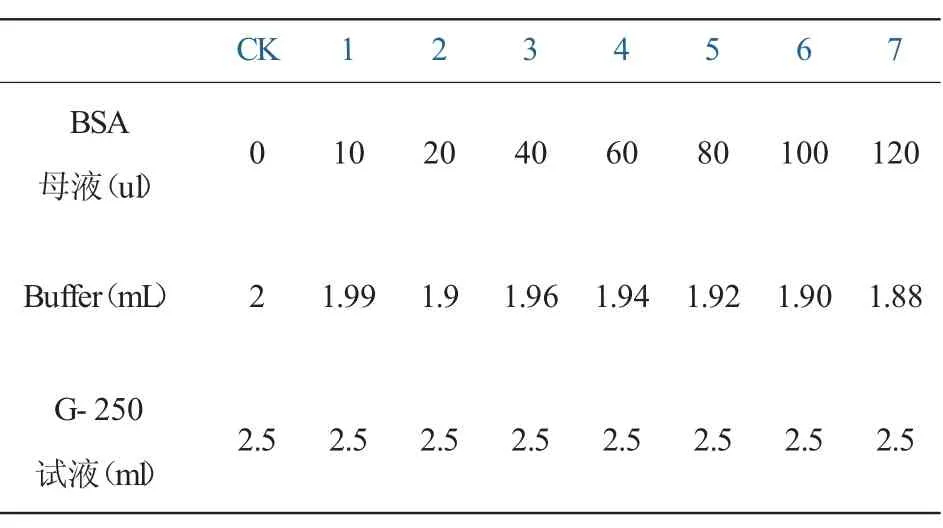
表2 蛋白质标准曲线应加试剂
4.样本测定。挑取田间麦蚜和敏感品系无翅健壮成蚜各300头,加入1 mLpH 7.0,0.04 M磷酸缓冲液匀浆,4℃,10 800 rpm离心15 min,取上清液0.05 mL,加入磷酸缓冲液0.95 mL,加入染色液5 mL,混合均匀后在2 min至1 h内测OD595值,由标准曲线计算得到麦蚜蛋白质含量。
5.乙酰胆碱酯酶活力测定。采用Gorun(1978)改进的Ellman方法,取四支试管,分别加入0.1 mL碘化硫代乙酰胆碱(ATCh)(10 mmol/L)作底物,前三支试管再分别加入0.1 mL AChE反应生成硫代胆碱和乙酸,在30℃水浴中反应15 min,振荡3次。然后加入0.9 mL显色剂(DTNB)显色并中止反应,第四支试管为空白管,在加入显色剂后补加0.1 mL酶液,将试剂振荡均匀,在412 nm下测定光密度(OD)值[17]。按照如下公式即可算出乙酰胆碱酯酶的比活力。
AChE的比活力比活力(mol/mg.Pr.min)=(ODV总)/(TPr. VE)
6.乙酰胆碱酯酶Km和Vmax的测定。Km是酶的特征性常数,该值的大小表明酶和底物之间亲和力的高低[21]。将麦蚜乙酰胆碱酯酶液稀释成所需倍数用于测定羧酸酯酶的Km及Vmax,将试管分成7组,每组设三个重复和一个对照,按照表3加入试剂[1]。
室温放置15 min,600 nm(或550 nm)下比色测定OD值。
根据蛋白含量的测定,计算出不同底物浓度下羧酸酯酶的比活力,然后根据Enzif软件计算出Km和Vmax值。
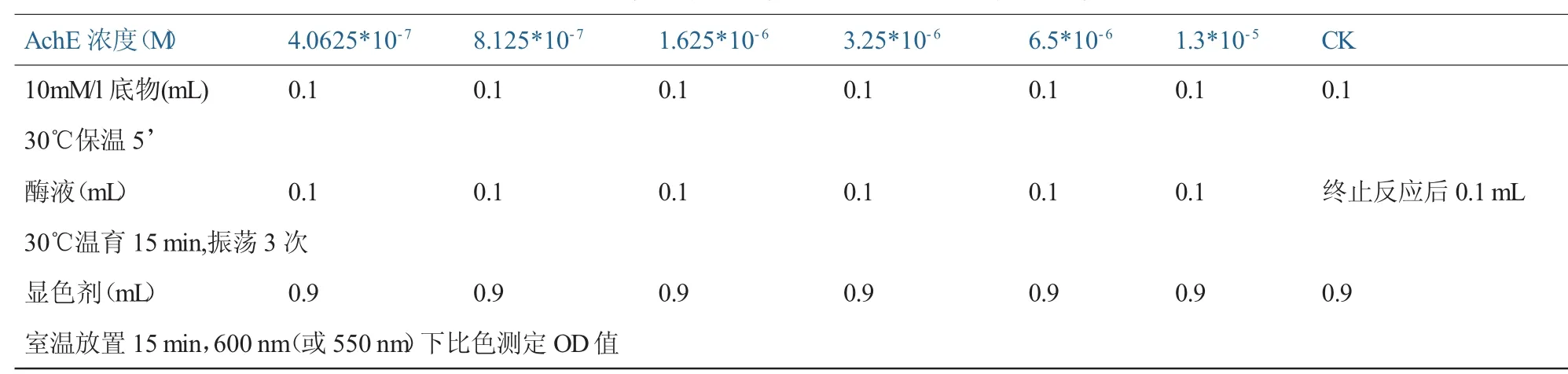
表3 乙酰胆碱酯酶Km和Vmax应加试剂
(三)羧酸酯酶活力测定
1.试剂配制。配制0.1 mg/mL的牛血清蛋白液及考马斯亮兰显色剂,0.03 Mα-NA,β-NA,α-NPr含0.03 M毒扁豆碱,磷酸缓冲液PBS(pH7.0,0.04 M);显色剂:1%固兰B盐+5%SDS(2:5)。
2.酶液制备。挑取田间麦蚜和敏感品系无翅健壮成蚜各300头,加入1 mLpH 7.0,0.04 M磷酸缓冲液匀浆,4℃,10 800 rpm离心15 min,上清液为酶液母液,将其分装置于-20℃,根据不同品系的抗性程度稀释20~30倍用于实验。
3.蛋白质标准曲线制作。将试管分成7组,每组设3个重复,按照表4加入试剂。

表4 羧酸酯酶活力测定应加试剂
混匀,室温放置10~20 min,分光光度计595 nm处测定吸光值,重复三次取平均值。以标准蛋白浓度(mL)为横坐标,OD值为纵坐标,做出蛋白质标准曲线。
4.麦蚜羧酸酯酶活力的测定。羧酸酯酶比活力测定方法参照高希武[22]的方法并稍加修改。取七支试管分别加入PH 7.0,0.04 mol/L磷酸缓冲液0.9 mL,3×10-4 M底物(含毒扁毒碱,1:1)1.8 mL,前六支试管中再分别加入适当稀释的酶液100 μL,30℃水浴中反应15 min后,加入0.9 mL显色剂(1%固蓝B盐:5%SDS2:5)终止反应。第七支试管为空白管,在加入显色剂后补加100 uL酶液。静置15 min后,在分光光度计中于600 nm(α-NA)或550 nm(β-NA)波长测定光吸收值。根据得到的OD值和酶液蛋白含量,按照如下公式即可算出羧酸酯酶的比活力。比活力(mol/mg.Pr.min)=(ODV总)/(TPr.VE)
其中:V总为反应总体系(mL);T为反应时间(min);Pr.为蛋白含量(mg/mL);VE为反应酶液体积(mL)。
5.麦蚜羧酸酯酶Km及Vmax测定。将麦蚜羧酸酯酶酶液稀释成所需倍数用于测定羧酸酯酶的Km及Vmax,将试管分成7组,每组设三个重复和一个对照,按照表5加入试剂。
室温放置15 min,600 nm(或550 nm)下比色测定OD值。
根据蛋白含量的测定,计算出不同底物浓度下羧酸酯酶的比活力,然后根据Enzif软件计算出Km和Vmax值。
(四)结果与分析
1.ACHE比活力测定。ACHE活力测定结果如表6,田间麦蚜AchE的比活力1.59±0.016 mol/mg.pr.min,室内麦蚜AchE的比活力为3.26±0.104 mol/mg.pr.min,室内麦蚜的AchE比活力是田间麦蚜的2倍。

表5 麦蚜羧酸酯酶Km及Vmax测定应加试剂

表6 田间麦蚜和室内品系乙酰胆碱酯酶比活力测定结果
2.ACHE的km值与Vmax。田间麦蚜的km值为25.76 mol/L大于室内麦蚜的20.4 mol/L,而最大反应速度为1.867 mol/mg.pr.min小于室内麦蚜的2.850mol/mg.pr.min。

表7 田间麦蚜和室内品系乙酰胆碱酯酶的Km与Vmax
3.羧酸酯酶比活力。当底物为α-NA时,田间麦蚜的比活力为48.65 mol/mg.Pr.min,小于室内麦蚜的121.11 mol/mg. Pr.min。而当底物为β-NA时,田间麦蚜的比活力为497.46± 8.50 mol/mg.Pr.min,大于室内麦蚜的348.11±1.29 mol/mg.Pr. min。

表8 田间麦蚜和室内品系羧酸酯酶的比活力
4.羧酸酯酶km值与Vmax。由实验结果表明,田间品系羧酸酯酶Km值均大于敏感品系(1.05~1.22倍);抗性品系羧酸酯酶最大反应速度Vmax也均大于敏感品系1.15倍左右。
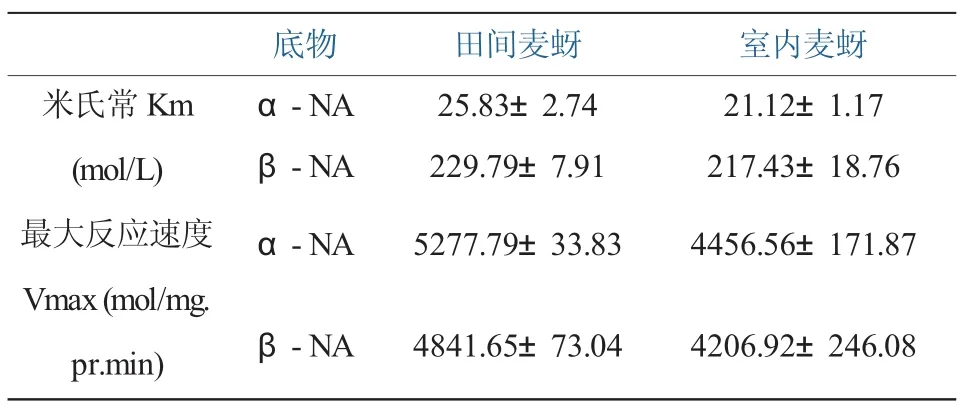
表9 室内麦蚜及田间麦蚜羧酸酯酶的Km和Vmax值
(五)讨论
害虫的抗药性问题是害虫综合治理中最重要的问题之一,明确害虫抗药性机制是害虫抗药性治理的基础。长期以来,人们一直关心对抗性生化机制的研究。近来,已有研究表明昆虫对杀虫剂的代谢能力提高是抗药性产生的重要原因,主要表现为一系列解毒酶活性的提高[23]。
乙酰胆碱酯酶是多种药剂的靶标酶,在正常昆虫中,杀虫剂使AchE活性降低,神经突触间的传递物质乙酰胆碱的正常释放或接收遭到破坏,打断神经传导,使昆虫死亡;而在抗性昆虫中,由于药剂的长期选择作用,突触间的物质传递活动已对药剂的干扰或破坏作用有了很强的适应性,发生了某些改变,甚至完全可以不受药剂的干扰而进行正常的神经传导活动,这样毒物药剂就失去了效用,昆虫不能因神经传导中断而死亡,害虫表现为抗药性。Torre,etal.(2002)曾指出,AChE是反映有机磷和氨基甲酸酯类农药污染最有效的生物标记酶.因此研究昆虫AChE的特性及有机磷和氨基甲酸酯类杀虫剂对其抑制作用,是研究这些杀虫剂的作用机理和抗性机理的基础。
本实验结果显示田间麦蚜的乙酰胆碱酯酶的活性与Vmax均小于室内品系,而田间麦蚜的Km值大于室内品系,Km是酶的特征性常数,该值的大小表明酶和底物之间亲和力的高低。可见田间麦蚜的AchE与底物的结合力较低,田间麦蚜的AChE对杀虫剂的敏感性下降,可能是田间麦蚜AChE的量过高所致,也可能是AChE性质结构发生了改变,这还有待进一步研究证明。
羧酸酯酶是昆虫体内重要的解毒酶系,它不仅具有广泛的底物特异性,可以水解羧酸酯键、酰胺键和硫酯键,在对含酯键杀虫剂的水解方面起着重要作用,而且还可以作为结合蛋白与杀虫剂分子不可逆结合,使其不能到达靶标,从而起到对杀虫剂的隔离作用。昆虫体内该酶活性的高低会直接影响其对杀虫药剂的抵抗能力。
本实验结果显示,当以α-NA为底物时,田间麦蚜的羧酸酯酶水解活性明显低于室内品系的,而动力学常数Km值与Vmax均大于室内品系;当以β-NA为底物时,田间麦蚜的羧酸酯酶水解活性,动力学常数Km,Vmax均与室内麦蚜无明显区别。
由上述结果可推断,田间麦蚜对于不同种类的农药产生高低不同的抗性与其体内的乙酰胆碱酯酶有显著关系而与羧酸酯酶关系不大。
四、结论
文章对田间麦蚜和实验室麦蚜进行了毒力测定以及生化测定。毒力测定使用了四大类别7种农药,结果显示,田间麦蚜相对于室内麦蚜来说,对于对于有机磷类的氧化乐果,菊酯类的敌杀死,氨基甲酸酯类的灭多威三种农药的LC50,田间麦蚜的LC50分别为敏感麦蚜的40倍、11倍、3倍。抗性明显。对于辛硫磷,毒死蜱,速灭杀丁和吡虫啉,田间麦蚜的敏感性较高,没有表现出抗性。由此可见,田间麦蚜对于某些农药
表现出明显抗性,该类农药应是经常用于麦蚜治理。
田间麦蚜对于生化测定来说,田间麦蚜的乙酰胆碱酯酶的活性及Vmax均小于室内品系,而Km值大于室内品系。对于其羧酸酯酶的水解活性,当以α-NA为底物时,田间麦蚜
的羧酸酯酶水解活性明显低于室内品系的,而动力学常数Km值与Vmax均大于室内品系;当以β-NA为底物时,田间麦蚜的羧酸酯酶水解活性,动力学常数Km,Vmax均与室内麦蚜无明显区别。这可能是由于本实验所用麦蚜均采自田间田间,对其用药背景不了解,其抗性背景较复杂.由上述结果可推断,田间麦蚜对于不同种类的农药产生高低不同的抗性与其体内的乙酰胆碱酯酶有显著关系而与羧酸酯酶关系不大。
本试验的结果只是初步研究了田间麦蚜的抗药性及抗性机制产生的原因.若要进一步了解乙酰胆碱酯酶是如何参与其抗性过程还需进一步做分子生物学的实验.
[1]潘怡欧.棉蚜对马拉硫磷的抗性机制研究[D].北京:农学与生物技术学院,2001.
[2]魏岑,黄绍宁.麦长管蚜的抗药性研究[J].昆虫学报,1988,31(2):148-156.
[3]高希武,周序国,王荣京,等.棉铃虫乙酰胆碱酯酶(ACNE)的体躯分布及部分纯化[J].昆虫学报,1998,41(增):19-25.
[4]姚洪渭,蒋彩英,叶恭银,等.白背飞虱乙酰胆碱与乙酞胆碱酯酶的体躯与亚细胞分布特征[J].浙江大学学报,2001,27(1):5-10.
[5]李腾武,高希武,郑炳宗,崔建州.小菜蛾不同亚细胞层羧酸酯酶的性质研究[J].农药学学报,1999,1(2):47-53.
[6]冯俊涛,王兴林,杨崇珍,等.陕西棉花苗蚜对几种杀虫剂的抗药性测定[J].西北农业大学学报,1996,24(3):63-67.
[7]韩召军,王荫长,尤子平.棉蚜对拟除虫菊酯类杀虫剂的抗性机理[J].南京农业大学学报,1995,18(3):54-59.
[8]韩召军,张明,王荫长.五种蚜虫羧酸酯酶的活力和同工酶谱变异的初步研究[J].南京农业大学学报,1987,4(增刊):31-35.
[9]李飞,韩召军,吴智锋,等.我国棉蚜抗药性研究现状[J].棉花学报,2001,13(2):121-124.
[10]洪波,关晓庆.宁夏地区麦蚜对几种杀虫剂的敏感性测定[J].宁夏农学院学报,2004,25(3):17-20.
[11]王晓军,陶美玲.麦长管蚜和禾谷缢管蚜对吡虫啉的抗药性研究[J].昆虫知识,2004,41(2):155-157.
[12]慕立义,王开运,姜家良,等.棉蚜对不同药剂抗性发生规律及其机理研究[J].杀虫药剂,1988,27(2):5-7.
[13]艾颖,邱星辉,何凤琴.棉蚜抗药性机理研究进展3[J].中国农业科学,2003,36(8):910-915.
[14]鲁艳辉,杨婷,高希武.禾谷缢管蚜和麦长管蚜玻璃管药膜法敏感毒力基线的建立[J].昆虫学报,2009,52(1):52-58.
[15]张莹,黄建,高希武.两种蜜蜂头部乙酰胆碱酯酶对杀虫药剂敏感度比较[J].农药学学报,2005,7(3):212-226.
[16]程桂林,刘润玺.新疆、田间棉蚜抗药性对比[J].农药,1997,36(11):6-9.
[17]郭晶,高菊芳,唐振华.羧酸酯酶及其在含酯类化合物代谢中的作用[J].农药,2007,46(6)8-10.
[18]王旭,高希武,郑炳宗,等.有机磷药剂对棉铃虫羧酸酯酶的抑制作用[J].昆虫学报,1998,41(增刊):12-14.
[19]高希武,胡慢华,郑炳宗.棉蚜不同抗性品系竣酸醋酶比较[J].植物保护学报,1997,24(4).
[20]相建华.麦蚜的发生特点与防治对策[J].现代农业科技,2007,11.
[21]李亮,付晓伟,郭线茹,等.几种药剂对2种麦蚜的室内毒力和田间药效[J].河南农业科学,2009,02:71-74.
[22]李腾武,沈福英,赵晓珍,等.不同种群小菜蛾乙酰胆碱酯酶性质研究[J].中国植保导刊,2005,4:12-13.
[23]魏绪强,钱坤,曾晓,等.羧酸酯酶与北京市不同城区德国小蠊对有机磷杀虫剂抗性的关系[J].中国媒介生物学及控制杂志,2008,19(5):1003-1004.
(编辑:张琼琼)
S481.4
B
1673-9019(2016)02-00-0
2016-01-06
李宏德(1990-),男,甘肃定西人,主要从事马铃薯良种生产工作。
猜你喜欢
特种经济动植物(2022年3期)2022-11-25
河北果树(2021年4期)2021-12-02
四川蚕业(2021年4期)2021-03-08
核农学报(2021年2期)2021-01-28
鞍山师范学院学报(2018年6期)2019-01-10
现代农业科技(2018年13期)2018-10-20
世界热带农业信息(2018年3期)2018-09-26
天然产物研究与开发(2018年8期)2018-09-10
天然产物研究与开发(2018年4期)2018-05-07
分析化学(2017年12期)2017-12-25
