
芦苇种群构件结构及生长分析*
2017-01-06 01:49宋金枝杨允菲
通化师范学院学报 2016年12期
宋金枝,韦 赫,杨允菲
(1.通化师范学院生命科学学院,吉林通化134001;2.吉林农业大学,吉林长春130024; 3.东北师范大学草地研究所,植被生态科学教育部重点实验室,吉林长春130024)
芦苇种群构件结构及生长分析*
宋金枝1,韦 赫2,杨允菲3
(1.通化师范学院生命科学学院,吉林通化134001;2.吉林农业大学,吉林长春130024; 3.东北师范大学草地研究所,植被生态科学教育部重点实验室,吉林长春130024)
利用大样本随机抽样的方法,采挖单个植株用来调查和测定,对芦苇种群构件及生长进行分析,结果表明,在成熟期,芦苇种群构件结构是由根、叶、杆、穗组成.芦苇各构件之间相关关系是极其显著的,株高与穗长之间用线性函数表示其相关程度最好,其决定系数R2为0.9462;芦苇株高及各构件生物量之间相关关系也是极其显著的,线性函数也是表现其相关程度最佳的函数模型,其决定系数R2为0.8345~0.9677.因此芦苇种群的生长规律是由线性函数表达出来,芦苇种群构件在生长期间呈现同速生长.
芦苇;构件;生长分析;同速生长
植物种群构件理论是植物种群生物学及生态学研究的一个新方向.构件是指存在多细胞结构的并互相联系结合的重复单元[1-2].在植物的发育生长期间,植物通过调节其构件的结构从而适应不断变化的环境;并且,每种植物构件在生长期间中都有它自己的规律性.通过研究植物构件的生长规律,能够知道植物发育生长期间各构件的生长状况和变化并对其进行定值描述,是深入研究植物适应与进化的基础[3-5].
芦苇(Phragmites communis)是根茎型禾草,营养繁殖力强,是典型的无性系植物,在合适环境条件下可形成单优群落,天然种群补充更新依靠根茎繁殖.芦苇一般在三月底发芽,五至六月份生长速度最快,七月中旬芦苇即将成熟,种子完全成熟一般在十月中下旬,收割期在霜降后.沼泽地、河漫滩和浅水湖都是芦苇喜生长的地方,常形成苇塘,全球各大洲都有分布.我国从北方到南方都有生长.在pH值6.5~9.0的偏酸、中、碱性,以及在滩涂盐土或水边也能生长发育.芦苇形成的种群通常是该地区的优势种或伴生种[6].芦苇主要在湿地环境中生长,芦苇茎秆笔直、高大、坚韧,纤维含量高,可作为原料应用在造纸工业中.因为通气组织遍布在芦苇的叶、叶鞘、茎和不定根中,所以它可以净化周围的污水,同时可减缓河流流速和促进泥沙下沉,因此芦苇种群在河流排污、排洪和护堤中起重要作用.有许多浅水湖芦苇湿地已被列入国际重要湿地名单[6].
近些年来,国内外关于芦苇的研究比较多,但对于芦苇植株生长规律的研究相对很少.本文以河北省文安县芦苇为研究对象,就芦苇株高、穗长、鲜叶重、杆鲜重、穗鲜重、叶干重、杆干重、穗干重等数量性状分别进行比较分析.从而得出该地区芦苇群构件结构特点和生长规律,为芦苇的种植和高效利用提供更为有效的理论支持.
1 研究地区与研究方法
1.1 研究地区
文安县地处河北省廊坊市文安县县域中心地带,位于华北平原相对低下部位,属于河流堆积地貌,平坦开阔,为多条河流下游.地势自西南向东北倾斜.文安镇受自然环境、太阳照射和季风影响,境内气温适中、热量充足,光照丰富、没有霜的时间多.形成了气候温暖,四季分明的气候特点.文安县季风气候明显,春秋两季处于过度季节,没有明显偏多风向,夏季多偏南风,冬季多偏北风.
1.2 研究方法
2015年10月中旬,即芦苇的成熟期,在河北省文安县区域芦苇生长的典型区域,采用大样本随机取样法进行取样.取样时,分别挖取32个大小不等的植株,去除根部泥土并编号.回到实验室有顺序的测定其株高、穗长、叶鲜重、杆鲜重、穗鲜重、叶干重、杆干重、穗干重.其中芦苇的各构件长度选用厘米尺测定,在测量时操作规范,认真记录数据并标好序号,注意不能破坏植株的各个构件,造成数据错误.芦苇的叶鲜重、杆鲜重以及穗鲜重采用 Sartorius BA210s型电子天平进行测定,然后将各构件装入袋中并标清编号,放进烘干箱中80℃恒温烘至恒重,然后再测定其生物量.
1.3 数据处理
选择Excel软件进行数据统计与分析,其生长分析的定量刻画模型采用线性函数、对数函数、指数函数、幂函数4种函数中相关性最高的函数模型来表示[7-9].
2 结果与分析
2.1 植株数量特征及构件的结构
芦苇的构件结构分为穗、杆、叶和根.统计分析表1表明的是32株芦苇各构件的数量特征,表1中的平均值表示的是芦苇种群的整体水平,标准差表示芦苇种群变异的绝对数量指标,变异系数反映样本变异的相对数量指标.最大值和最小值体现出芦苇种群的实际范畴[10].株高作为芦苇植株间最显著的构件特征,在一定水平上反映了该地区芦苇生长状态.芦苇植株的各个构件之间具有一定的相关性,从本次分析结果可以看出该地区比较适合芦苇生长[8].由表1可知,株高的最大值与最小值相差2.35倍,变异系数是21.32%;穗长的最大值与最小值相差5.5倍,变异系数是27.91%,说明芦苇株高变异幅度没有穗长的变异幅度大.通过比较芦苇的叶、杆、穗的干鲜重,可以看出芦苇的杆构成了芦苇种群构件的主体.芦苇株高在77.51cm~182.31cm之间,平均株高130.75cm,变异系数最小,说明芦苇的株高是集中在均值附近而不是离散的.穗干重的变异系数最大82.22%,表明穗干重的变异幅度最大.
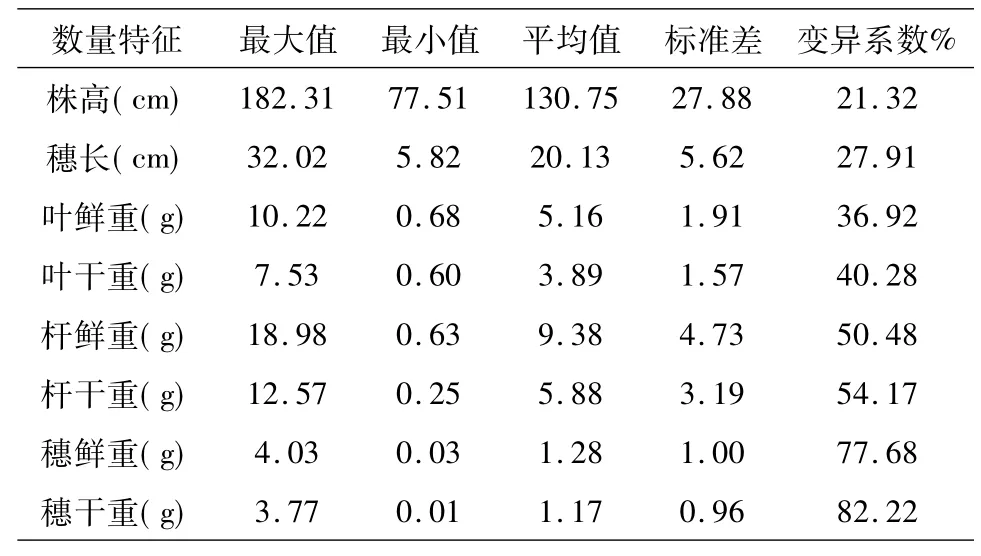
表1 芦苇构件的数量特征(n=32)
2.2 芦苇的株高、穗长之间相关性分析
通过分析计算,芦苇株高、穗长之间相关关系极其显著,用线性函数来表示其相关程度最好(图1),其决定系数R2是0.9462,揭示了株高和穗长的关系,绝大部分还是受遗传因素的影响比较大,受环境的影响比较小,但具备一定的生态可塑性.通过线性函数可知,株高和穗长之间呈现同速生长规律,所以随着株高的增加,穗长以同样比例的线性函数形式增加;通过图1的拟合方程(y=kx+b)可以知道,即芦苇的株高达到28.06cm(y=0)以上时才能长出穗.
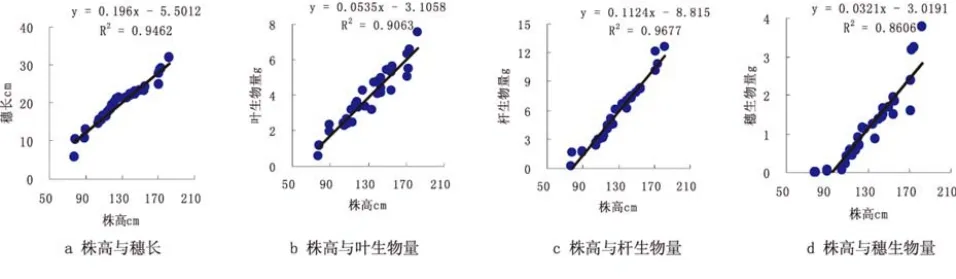
图1 芦苇的株高与穗长、叶生物量、杆生物量、穗生物量的观测值及拟合曲线

图2 芦苇叶生物量、杆生物量、穗生物量之间的观测值及拟合曲线
2.3 芦苇的株高及各构件生物量之间相关性分析
通过分析计算,芦苇各构件及株高之间相关关系极显著,用线性函数来表示其相关程度最好图(图1 b~d),其决定系数R2在0.8345~0.909之间.芦苇的株高与各构件的生物量之间的函数关系说明了,随着株高高度的增长,各构件生物量以特定的比例增长,其决定系数 R2分别为 0.9063、0.9677、0.8606,表明株高与各构件的生物量之间的关系主要是由物种自身遗传所决定,与外部环境的关系不大.换句话说,随着株高高度的增高,叶生物量的90.63%是以线性函数形式增长的,随着株高高度的增高,杆生物量的96.77%是以线性函数形式增长的,随着株高高度的增高,穗生物量的86.06%是以线性函数形式增长的;芦苇构件叶生物量、杆生物量、穗生物量之间也有密切的联系,均用线性函数来表示其相关程度最好图(图2 e~g),决定系数R2>0.8345,体现出各构件生物量拥有同速生长的特点,同时环境因素对芦苇生长的改变很小.
3 讨论
芦苇广泛分布于沼泽地、河漫滩和浅水湖.在成熟期,芦苇种群构件由根、杆、叶和穗组成.在整体水平上,芦苇杆占的比例最大,构成了芦苇种群构件的主体.
变异系数是衡量数据中各观测值变异程度的一个统计量,表示数据的离散程度,如果几组数据的测量尺度相差太大,直接使用标准差来进行比较不合适,而变异系数可以做到这一点,计算变异系数的公式为:变异系数=(标准差/平均值)×100%.通常情况下变异系数大于100%属于强变异,超过10%且小于100%的属于中等变异,不到10%的属于弱变异.根据芦苇构件分析数据可知,芦苇的株高、穗长以及叶、杆、穗的鲜重与干重变异均算作中等变异.
在植物生长发育期间,构件形态变化有两种模式,一种是同速生长模型,另一种是异速生长模型.同速生长型常以直线函数形式表现数量性状间的关系,而异速生长型则常以曲线函数来表示[11-15].通过本次研究可以得出,芦苇构件在发育生长期间,呈现为同速生长规律.
在研究植物各构件的生长分析中,R2表示拟合方程的决定系数,R2值的大小决定了相关的密切程度,因此可用来研究遗传因素所占的比重,而1-R2则表现为外部环境所占的多少[16].通过本次对芦苇种群构件结构及生长分析的研究中,可发现,决定系数R2为0.8345到0.9677之间,所以说芦苇各构件的发育生长,受到遗传因素的作用比较明显,而芦苇受环境因素的作用程度比较小,但芦苇种群还有相当程度的生态可塑性[17].
[1]Francisco L,Sara GF,José MS,et al.Analysing hierarchically -structured fitness and modular dynamics in plants:Integration of conceptsfrom population dynamics[J].Perspectives in Plant Ecology Evolutionand Systematics,2002(5):123-129.
[2]Harper J L,White J.The demography of plants[J].Annual Review of Ecology and Systematics,2003,5(1):419-463.
[3]黎云祥,刘玉成,钟章成.植物种群生态学中的构件理论[J].生态学杂志,1995,4(6):35-41.
[4]刘庆,钟章成.无性系植物种群生态学研究进展及有关概念[J].生态学杂志,1995,4(3):40-45.
[5]宋金枝,杨允菲,李海燕,等.长白山区垂穗披碱草无性系结构与生长分析[J].江苏农业科学,2013,41(1):327-329.
[6]杨允菲,郎惠卿.不同生态条件下芦苇无性系种群调节分析[J].草业学报,1998,7(2):1-9.
[7]田迅,杨允菲.西辽河平原不同生境草芦种群分株生长的可塑性[J].草地学报,2004,12(1):17-20.
[8]李博,刘存歧,闫蓬勃,等.白洋淀湿地芦苇生长参数的线性分析[J].湖北农业学报,2010,2(49):316-318.
[9]宋金枝,杨允菲,董雪,等.长白山区月见草种群构件结构及生长分析[J].通化师范学院学报,2015,5(36):11-12.
[10]宋金枝,李海燕,周丽威.长白山区华北翦股颖无性系构件的结构及生长[J].生态学杂志,2011,30(10):2145-2148.
[11]王俊炜,李海燕,杨允菲.温带地区4种园林灌木叶片的生长规律[J].东北师范大学学报,2005,37(1):95-98.
[12]杨允菲,张宝田.松嫩平原人工草地野大麦无性系构件生物量的结构及生产规律[J].应用生态学报,2004,15(8):1378-1382.
[13]王立,周婵,杨允菲.林间草地寸草苔种群的生态可塑性分析[J].中国草地,2001,23(4):6-10.
[14]钟章成,曾波.植物种群生态研究进展[J].西南师范大学学报,2001,26(2):230-236.
[15]宋金枝,王蕾,杨允菲.大苞萱草种群无性系结构与生长分析[J].江苏农业科学,2013,41(5):153-154.
[16]杨允菲,李建东.松嫩平原全叶马兰种群不同龄级分株的生长策略[J].应用生态学报,2003,14(12):2171-2175.
[17]宋金枝,杨允菲.长白山区南部黄刺玫花构件的结构及生长分析[J].东北林业大学学报,2008,36(9):21-23.
(责任编辑:陈衍峰)
Q945.3
A
:1008-7974(2016)06-0023-03
10.13877/j.cnki.cn22-1284.2016.12.007
2016-10-20
国家自然科学基金“东北草原碱斑极端生境两种广布无性系禾草的趋异适应机理”(31472134)
宋金枝,女,吉林梅河口人,教授.
杨允菲,E-mail:yangyf@nenu.edu.cn
猜你喜欢
麦类作物学报(2022年7期)2022-08-09
海外文摘·文学版(2022年4期)2022-04-14
黄河之声(2021年19期)2021-02-24
农村百事通(2019年17期)2019-10-08
农村百事通(2018年16期)2018-09-29
岁月(2018年2期)2018-02-28
中国诗歌(2017年12期)2017-11-15
现代农业科技(2017年1期)2017-03-06
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
