
拟南芥苯丙氨酸解氨酶基因的克隆与表达分析
2017-01-06 02:21赵丹
山西农业科学 2016年12期
赵丹
(山西农业大学信息学院,山西太谷030800)
拟南芥苯丙氨酸解氨酶基因的克隆与表达分析
赵丹
(山西农业大学信息学院,山西太谷030800)
苯丙烷途径是植物体内广泛存在的次生代谢途径,此代谢过程中的第1个关键酶为苯丙氨酸解氨酶(Phenylalanine ammonia-1yase,PAL)。苯丙氨酸解氨酶通过对基因的起始转录、频率的控制和对外界诱导因子的响应来调控基因的表达,并且其表达水平对于下游黄酮类物质的合成起着非常重要的作用。在一定条件下,苯丙氨酸解氨酶可利用其特殊的酶学性质逆向催化生产L-苯丙氨酸。通过一系列的生物信息学分析,确定拟南芥中苯丙氨酸解氨酶的存在,并进行基因的克隆及表达分析,为L-苯丙氨酸的生产提供新的物种来源。
拟南芥;苯丙氨酸解氨酶;生物信息学;克隆
苯丙氨酸解氨酶是1961年在大麦中发现的[1],其是苯丙烷途径的第1个关键酶,在各种各样的植物中均有存在,而且在少量微生物中也有存在。苯丙氨酸解氨酶是一种将苯丙氨酸转化为酚类化合物的重要蛋白酶,决定了酚类化合物合成的速率[2]。同时,苯丙氨酸解氨酶通过对基因的起始转录、频率的控制以及对外界诱导因子的响应来调控基因的表达,并且其表达水平对于下游黄酮类物质的合成起着非常重要的作用。在酸碱值为10,过量的铵根离子和酚类化合物同时存在的条件下,苯丙氨酸解氨酶可逆向催化酚类化合物加氨生成L-苯丙氨酸[3-5],这种特殊的酶学性质被广泛地应用在生物化工领域,以创造更高的价值。
提高L-苯丙氨酸产量的一个重要方法是在众多的苯丙氨酸解氨酶源中筛选出一种活力较高并且稳定性好的物种[6-8]。拟南芥的生长过程较短,培养条件也相对较低,便于大量培育,是分子遗传性与农业研究中良好的材料[9]。同时,由于拟南芥基因高度纯合的特点,使用不同的物理因素或者化学因素对拟南芥进行处理时其容易产生突变,生成突变植株,便于对其进行代谢功能及调控酶的研究[10-11]。建立拟南芥中苯丙氨酸解氨酶的研究机制,为L-苯丙氨酸的生产提供新的物种来源,对于我国的农业研究和农作物的发展具有重要的影响及意义。
1 材料和方法
1.1 材料
供试材料为拟南芥幼苗。
1.2 试剂及仪器
1.2.1 试剂PCR引物合成于生工公司;反转录试剂盒、TE缓冲液购于TAKARA公司;苯酚、氯仿、戊酮、异丙醇无水乙醇试剂均为分析纯。
1.2.2 主要仪器培养箱、PCR仪、-20℃冰箱、冷冻离心机、灭菌锅、电子天平、pH计、电泳仪、凝胶成像系统。
1.2.3 主要试剂配制提取缓冲液:Rris-HCl,EDTA,NaCl,SDS;氯仿-戊酮-乙醇体积比为80∶4∶16。
1.3 方法
1.3.1 拟南芥苯丙氨酸解氨酶序列查找在NCBI中通过苦荞苯丙氨酸解氨酶的BLAST分析,查找可能的拟南芥苯丙氨酸解氨酶序列。其他物种的苯丙氨酸解氨酶序列从NCBI蛋白数据库中获得。
1.3.2 生物信息在线分析网站http://swissmodel. expasy.org/Swiss-model在线数据库;http://www.ncbi. nlm.nih.gov/NCBI在线数据库;http://prosite.expasy. org/PROSITE在线数据库;http://www.expasy.org/Expasy在线数据库;https://npsa-prabi.ibcp.fr/cgi-bin/ NPS@在线数据库。
1.3.3 生物信息分析软件预测蛋白跨膜结构利用TMHMM软件;信号肽预测工具利用SignalP软件;亚细胞定位分析利用WOLFPSPORT软件;蛋白同源性比对利用NCBI中的BLASTX程序;蛋白序列比对分析利用DNA MAN软件;蛋白理化性质分析利用Protparam工具。
1.3.4 拟南芥基因组的快速提取在50 mL离心管中加入20 mL提取缓冲液,50℃水浴预热;取拟南芥幼苗5~15 g,粉碎;加入25 mL氯仿-戊酮-乙醇溶液,颠倒混匀;在室温下5 000 r/min离心5 min;移取上清液放置50 mL离心管中,加入1倍体积异丙醇,摇动混匀,在室温下静置,出现絮状DNA沉淀;在1.5 mL离心管中加入1 mL TE,用玻璃棒取出DNA絮团,用吸水纸吸干水分,再加入含TE的离心管中,使DNA溶解于TE中。
1.3.5 拟南芥总RNA的提取收取拟南芥放在灭过菌的研钵中,加入液氮捣碎,迅速转入1.5 mL离心管中,以3 000 r/min离心2 min,弃上清,将沉淀弹起;加入1 mL的Trizol试剂于1.5 mL离心管中,轻摇混匀,放置5 min;加入200 mL的氯仿到离心管中,用手剧烈摇晃15 s,在室温下放置10 min,管内溶液分层,于4℃,12 000 r/min离心15 min;将上层水相转移到干净的1.5 mL离心管中后,吸取等体积的异丙醇加入离心管中,上下颠倒摇晃至混匀,室温下放置10 min,4℃12 000 r/min离心15 min,此时可见RNA沉淀;弃上清,加入用RNA酶无菌水配制的75%乙醇,上下颠倒摇晃洗涤沉淀,4℃12 000 r/min离心10 min;弃上清,无菌条件下干燥5 min,将RNA溶于30 μL的去RNA酶无菌水中。
1.3.6 拟南芥总cDNA的合成参照TAKARA公司的反转录试剂盒,以提取到的总RNA为模板反转录为cDNA,反转录扩增体系为37℃反转录15 min,85℃5 s终止反应,产物4℃保存(表1)。

表1 拟南芥总cDNA的合成
1.3.7 拟南芥中PAL基因的克隆以拟南芥cDNA为模板,利用PCR技术扩增PAL基因的开放阅读框,并使用半定量方式检验PAL基因在不同时期拟南芥幼苗中的表达情况(表2)。
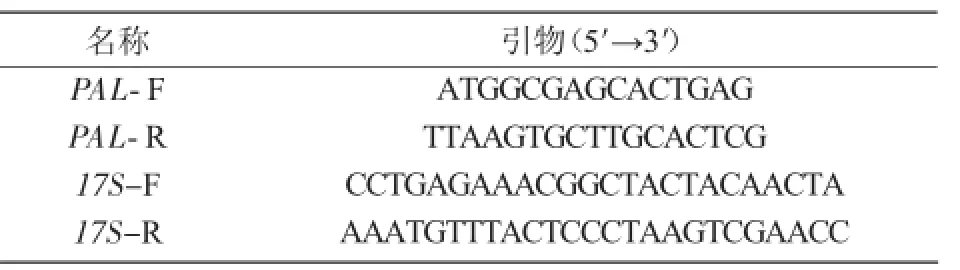
表2 试验所用引物
2 结果与分析
2.1 拟南芥苯丙氨酸解氨酶序列
在NCBI中通过苦荞苯丙氨酸解氨酶的BLAST分析,查找可能的拟南苯丙氨酸解氨酶序列,其结果如下。
MEINGAHKSNGGGVDAMLCGGDIKTKNMVIN AEDPLNWGAAAEQMKGSHLDEVKRMVAEFRKPV VNLGGETLTIGQVAAISTIGNSVKVELSETARAGVN ASSDWVMESMNKGTDSYGVTTGFGATSHRRTKNG VALQKELIRFLNAGIFGSTKETSHTLPHSATRAAML VRINTLLQGFSGIRFEILEAITSFLNNNITPSLPLRGTI TASGDLVPLSYIAGLLTGRPNSKATGPNGEALTAEE AFKLAGISSGFFDLQPKEGLALVNGTAVGSGMASM VLFETNVLSVLAEILSAVFAEVMSGKPEFTDHLTHR LKHHPGQIEAAAVMEHILDGSSYMKLAQKLHEMD PLQKPKQDRYALRTSPQWLGPQIEVIRYATKSIERE INSVNDNPLIDVSRNKAIHGGNFQGTPIGVSMDNTR LAIRAIGKLMFAQFSELVNDFYNNGLPSNLTASRNP SLDYGFKGAEIAMASSIEREINSVNDNPLIDVSRNK AIHGGNFQGTPIGVSMDNTRLAIRAIGKLMFAQFSELVNDFYNNGLPSNLTASRNPSLDYGFKGAEIAMAS YCSELQYLANPVTSHVQSAEQHNQDVNSLGLISSR KTSEAVDILKLMSTTFLVAICQAVDLRHLEENLRQT VKNTVSQVAKKVLTTGVNGELHPSRFCEKDLLKV VDREQVYTYADDPCSATYPLIQKLRQVIVDHALVN GESEKNAVTSIFHKIGAFEEELKAVLPKEVEAARA AYDNGTSAIPNRIKECRSYPLYRFVREELGTELLTG EKVTSPGEEFDKVFTAICEGKIIDPMMECLNEWNG APIPIC
2.2 蛋白质的理化分析
为了更加准确地了解拟南芥苯丙氨酸解氨酶,研究其理化性质是必不可少的,理化性质更能直接简单地为植物学研究提供很大的帮助。利用生物信息分析软件Protparam对拟南芥苯丙氨酸解氨酶进行理化分析,其结果如表3所示。
由表3可知,拟南芥苯丙氨酸解氨酶是由725个氨基酸组成的蛋白质,其分子量为78 782.7 Da,等电点PI为5.96。
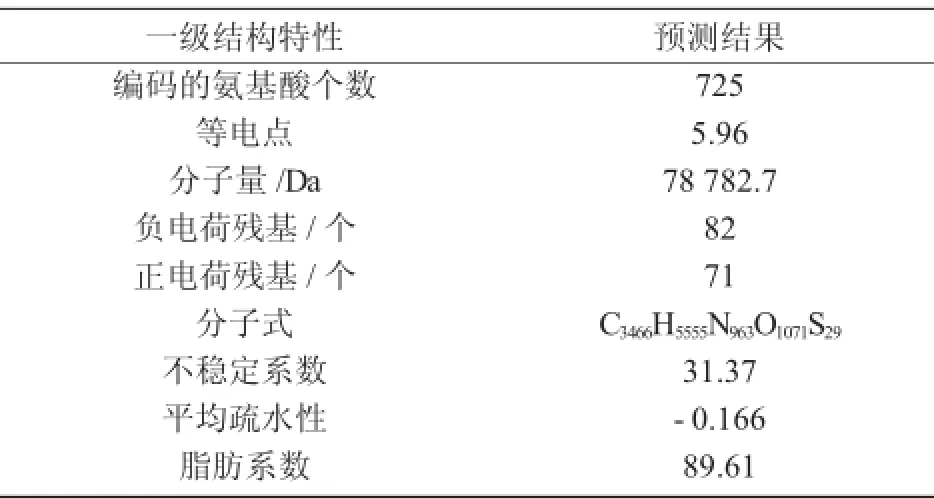
表3 蛋白质的理化分析
2.3 蛋白的功能域及保守域分析
使用PROSITE数据库[12]对拟南芥苯丙氨酸解氨酶氨基酸序列进行功能域和功能位点分析,以确定其是否具有保守功能域及其功能域的具体位置(图1)。由图1可知,拟南芥苯丙氨酸解氨酶氨基酸序列在第207~223位点存在一段保守功能域,序列为GTITASGDLVPLSYIAG,而此序列为丙氨酸解氨酶家族蛋白的特征序列,证明查找的蛋白序列存在苯丙氨酸解氨酶的功能基础。

2.4 蛋白质疏水性预测
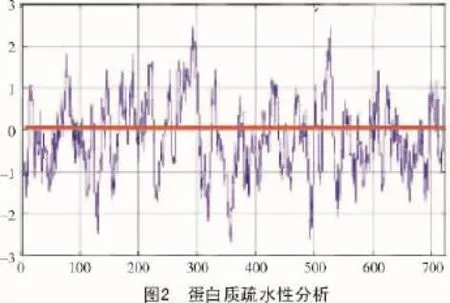
采用ProtScale(http://ca.expasy.org/tools/protsca le.html)软件[13]对拟南芥苯丙氨酸解氨酶进行疏水性分析(图2)。图2横轴表示的是氨基酸的位置,纵轴代表的是氨基酸平均疏水指标,并且图中的正值和负值分别表示的是疏水性和亲水性;由图2可知,较粗的横线为亲疏水性零点,此蛋白序列在亲疏水性正负两侧分布均匀,结合蛋白质一级结构分析可以得出,此蛋白总体没有表现出明显的亲水性或疏水性,符合苯丙氨酸解氨酶的特性。
2.5 蛋白跨膜结构预测
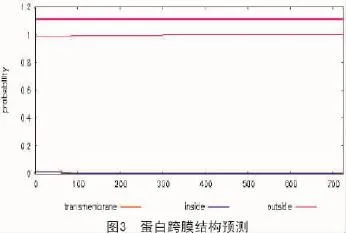
本试验利用TMHMM在线生物信息分析网站[14]中专业预测蛋白跨膜结构的软件,对拟南芥苯丙氨酸解氨酶进行蛋白跨膜结构预测(图3)。图3横轴表示的是氨基酸的位置,纵轴代表的是跨膜结构的可能性,图3结果显示,拟南芥苯丙氨酸解氨酶所有位点均没有存在跨膜结构的可能性。结合跨膜结构预测结果以及蛋白质疏水性预测结果表明该蛋白亚单位不含有跨膜结构域。
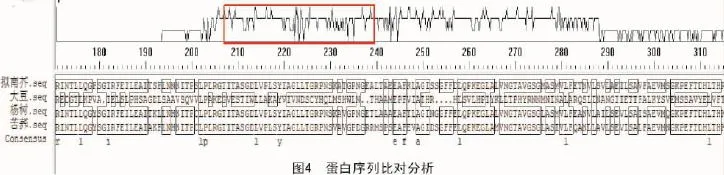
2.6 蛋白序列比对分析
通过NCBI查找发现,拟南芥中苯丙氨酸解氨酶有725个氨基酸,用DNA MAN软件[15]把其氨基酸的序列和大豆、杨树、苦荞中的苯丙氨酸解氨酶蛋白序列进行对比,结果显示,不同生物物种的苯丙氨酸解氨酶蛋白序列存在一定的序列一致性,并存在一个较保守的结构域,主要集中在210~230个氨基酸之间(图4)。这样的序列一致性符合苯丙氨酸解氨酶保守功能域的特性,证明拟南芥中苯丙氨酸解氨酶的结构域是保守的,能够执行相应的生物学功能,具有和其他物种苯丙氨酸解氨酶相似的生理意义。
2.7 蛋白质的三级结构分析
蛋白质的三级结构是蛋白质在二级结构的基础上进一步组成的,它具有疏水核位于内部、亲水链位于内部的特点[16-17]。利用Swiss-model在线数据库[18](http://swissmodel.expasy.org/)对拟南芥苯丙氨酸解氨酶进行三级结构预测,其结果如图5所示。
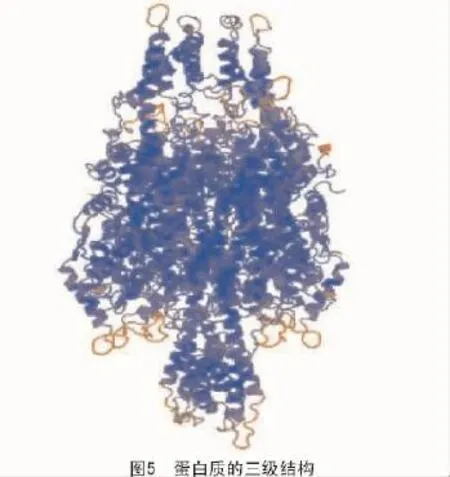
从图5可以看出,拟南芥苯丙氨酸解氨酶主要由α-螺旋组成,结构十分稳定。而α-螺旋在DNA结合基序(DNA binding motifs)中有非常重要的作用,比如在锌指结构、亮氨酸拉链、螺旋-转角-螺旋等基序中都含有α-螺旋。拟南芥苯丙氨酸解氨酶含有丰富的α-螺旋结构,其可能指导苯丙氨酸解氨酶对相关基因的起始转录、频率的控制和对外界诱导因子的响应,从而调控黄酮类物质的合成。
2.8 拟南芥中苯丙氨酸解氨酶基因序列的克隆
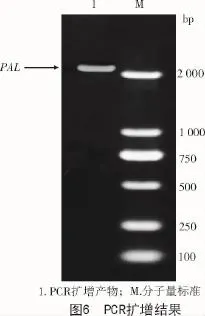
上述的试验验证了通过生物信息学方式查找到的拟南芥中苯丙氨酸解氨酶具有保守的功能结构域,是具有生物功能和意义的活性蛋白。为了进一步研究拟南芥中苯丙氨酸解氨酶的功能,进行基因过表达或转基因操作,以PAL-F与PAL-R为引物(表2),以拟南芥cDNA作为模板,PCR扩增出约2 178 bp的PAL基因片段(图6)。
2.9 拟南芥中苯丙氨酸解氨酶基因的表达分析
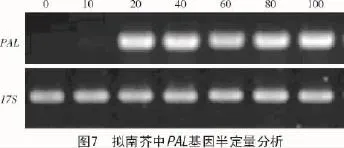
取拟南芥第0,10,20,40,60,80,100天的苗株进行qRT-PCR分析(图7),以17S为对照,结果表明,拟南芥中苯丙氨酸解氨酶基因在幼苗前20 d表达量不高,20 d后开始大量表达,并保持稳定的表达趋势,这可能是由次生代谢的开始所引起的。证明在拟南芥中苯丙氨酸解氨酶能够稳定存在,为L-苯丙氨酸的生产提供了可能的新的物种来源。
3 结论
生物信息技术的使用使我们对庞大基因库的分析变得更加高效、快速,这种分析方法可以使日后的研究实现“事半功倍”的效果,能够为之后试验研究提供重要线索,提升了日常工作效率[19-20]。本研究运用生物信息学分析软件从结构组成、基本理化性质、疏水性/亲水性、三级结构以及功能域等方面对拟南芥苯丙氨酸解氨酶进行预测和分析,结果表明,拟南芥PAL基因是由2 628个碱基所组成,其所编码含有725个氨基酸残基的多肽序列,蛋白分子量为78 782.7 Da;氨基酸中大部分没有显现出明显的亲水性或疏水性,并且等电点PI为5.96;该蛋白不可以进行跨膜运动,不含有跨膜结构域。对拟南芥苯丙氨酸解氨酶氨基酸序列进行功能域和功能位点分析,结果发现,其含有保守结构域。基因的克隆和表达谱分析表明,拟南芥中苯丙氨酸解氨酶基因在幼苗前20 d表达量不高,20 d后开始大量表达,并保持稳定的表达趋势,这可能是由于次生代谢的开始所引起的。证明在拟南芥中苯丙氨酸解氨酶能够稳定存在,为L-苯丙氨酸的生产提供了可能的新的物种来源。
[1]蔡朱男,余应年,罗建红.水稻苯丙氨酸氨解酶基因在大肠杆菌BL21DE3中的表达[J].中国病理生理杂志,2000(1):12-16.
[2]江柯.黏红酵母CIBASA1401苯丙氨酸解氨酶的分离纯化与性质研究[D].成都:四川大学,2004.
[3]李冀新,张超.L-苯丙氨酸的生产及研究进展[J].氨基酸和生物资源,2006(2):51-54.
[4]刘卫红,王燕,杜何为.银杏叶苯丙氨酸解氨酶分离纯化条件的研究[J].湖北农学院学报,2004(3):196-198.
[5]缪元颖,李化,杨亚力.苯丙氨酸解氨酶高活性红酵母(Rhodotorula sp.)CIBASA1401菌株的鉴定[J].应用与环境生物学报,2004(2):242-245.
[6]荣瑞芬,郭堃,厉重先.紫外照射诱导采后番茄苯丙氨酸解氨酶的分离纯化[J].北方园艺,2007(12):1-4.
[7]欧阳光察,应初衍,沃少根.植物苯丙氨酸解氨酶的研究:Ⅵ.水稻、小麦PAL的纯化及基本特性[J].植物生理与分子生物学学报,1985(2):204-214.
[8]董艳珍.植物苯丙氨酸解氨酶基因的研究进展[J].生物技术通报,2006(S1):21-24.
[9]马立安,江涛,张忠明.拟南芥Ran2基因的原核表达及产物的纯化[J].长江大学学报:自科版农学卷,2007(1):4-6.
[10]宋柬,马会勤,郝佳.大豆苯丙氨酸解氨酶在大肠杆菌中的重组表达及活性鉴定[J].食品科学,2006(7):29-35.
[11]徐虹,欧阳平凯.深红酵母转化反式肉桂酸生成L-苯丙氨酸的研究[J].工业微生物,2000(2):30-33.
[12]Havir E A.Phenylalanine ammonia-lyase:Purification and characterization from soybean cell suspension cultures[J].Archives of Biochemistryand Biophysics,1981(2):556-563.
[13]孙亚莉,郝辉芳,郭梦瑶.水稻耐盐基因ZFP1及蛋白的生物信息学分析生物技术[J].山西农业科学,2015,43(12):1585-1588,1616.
[14]Zhang X F,Yang F Y.Further study on the role of Mg2+in lipid-protein interaction of the reconstituted porcine heart mitochondrial H+-ATPase[J].Biochim Biophys Acta,1989,976:53-62.
[15]王东,杨金水.棉花类耐盐锌指蛋白基因的克隆与结构分析[J].复旦学报:自然科学版,2002(1):46-50.
[16]邹蔚然,郭万株.蛋白激酶基因的克隆和生物信息学分析[D].成都:四川农业大学,2008:21-26.
[17]Matus J T,Loyola R,Vega A,et al.Post-veraison sunlight exposure induces MYB-mediated transcriptional regulation of anthocyanin and flavonol synthesis in berryskins ofVitis vinifera[J].Journal of Experimental Botany,2009,60(3):853-867.
[18]王剑利,杨章民.利用生物信息学技术研究蛋白功能的几种方法[J].国外医学:分子生物学分册,2001,23(4):255-256.
[19]陈润生.生物信息学[J].生物物理学报,1999,15(1):5-6.
[20]李松,王英.生物信息学在生命科学研究中的应用[J].热带医学杂志,2009(10):1218-1221.
Cloning and Expression Analysis of Phenylalanine Ammonia-lyase Gene fromArabidopsis thaliana
ZHAODan
(College ofInformation,Shanxi Agricultural University,Taigu 030800,China)
Phenylalanine ammonia-lyase(PAL)is the first key enzyme involved in the metabolism of phenylpropanoid,which is a widespread metabolic pathwayin plants.Phenylalanine ammonia-lyase(PAL)regulates the expression ofgenes bytranscription initiation, frequency control and response to external factors,and its expression level plays an important role in downstream flavonoid synthesis. Under certain conditions,phenylalanine ammonia-lyase can be used to reverse-catalyze the production ofL-phenylalanine by its special enzymatic properties.In this paper,the presence of phenylalanine ammonia-lyase in Arabidopsis thaliana was determined by a series of bioinformatics analysis,and cloning and expression of the gene analysis,for the production of L-phenylalanine to provide a newspecies source.
Arabidopsis thaliana;phenylalanine ammonia-lyase;bioinformatics;clone
Q943.2
A
1002-2481(2016)12-1767-05
10.3969/j.issn.1002-2481.2016.12.07
2016-10-27
赵丹(1986-),男,山西太原人,助教,主要从事生物化学与分子生物学研究工作。
猜你喜欢
世界最新医学信息文摘(2021年24期)2021-04-29
科技视界(2020年26期)2020-09-24
科技视界(2020年17期)2020-07-30
国际放射医学核医学杂志(2020年4期)2020-07-27
生命科学研究(2018年1期)2018-05-29
化学教学(2018年1期)2018-02-28
上海农业学报(2017年3期)2017-04-10
食品工业科技(2015年11期)2015-05-05
河北科技大学学报(2015年6期)2015-03-11
植物营养与肥料学报(2014年1期)2014-03-11
