
氮钾配施对甘薯光合产物积累及分配的影响
2017-07-31 23:47汪顺义刘庆史衍玺李欢
中国农业科学 2017年14期
汪顺义,刘庆,史衍玺,李欢
氮钾配施对甘薯光合产物积累及分配的影响
汪顺义,刘庆,史衍玺,李欢
(青岛农业大学资源与环境学院,山东青岛266109)
【目的】探讨氮钾配施对甘薯光合产物转移分配的影响及其生理机制。【方法】开展两年田间试验,设CK(不施肥)、单独施氮(75 kg N·hm-2)、单独施钾(150 kg K2O·hm-2)和氮钾配施(75 kg N·hm-2+ 150 kg K2O·hm-2)4个处理,在生长前期(40 d)和薯块膨大期(100 d)分别进行13C叶片标记。测定了功能叶13C积累量和分配率、蔗糖合成酶(SS)和磷酸蔗糖合成酶(SPS)活性、叶绿素荧光和光合特性、干物质积累和产量等指标,并进行逐步回归分析、通径分析和RDA分析。【结果】与单独施氮和单独施钾相比,氮钾配施处理2014和2015年分别增产7.9%—10.1%和9.3%—10.7%。双因素分析表明,氮钾配施对甘薯产量的增加呈显著的正交互效应,其中,2014年交互效应值为0.95 t·hm-2,2015年交互效应值为1.35 t·hm-2。在生长前期和薯块膨大期,与氮、钾处理相比,氮钾配施处理显著提高了甘薯功能叶光合及叶绿素荧光特性,从而促进了两关键生长期光合产物的积累。其中CO2同化速率对应的量子产额(ΦCO2)提高27.1%—39.7%,净光合速率(n)提高9.1%—20.2%,甘薯地上部和地下部13C总积累量提高26.3%—42.2%。氮钾配施条件下光合产物分配在两生长期内存在差异。生长前期,氮钾配施处理通过提高叶片SS和SPS酶活性,显著提高了地上部13C分配率(达60.7%)(<0.05),促进光合产物在源器官分配;薯块膨大期,氮钾配施处理显著提高块根中SS和SPS酶活性,光合产物在库-源器官膨压差的作用下由地上部向地下部转运,显著提高13C向块根分配(13C分配率为71.6%,<0.05)。逐步回归分析表明,SS和SPS酶活性、光合特性和叶绿素荧光特性是调控甘薯光合产物分配的关键指标(1=0.954,2=0.912);通径分析表明,生长前期氮钾配施对甘薯13C分配的影响直接作用系数较大的是n、/和SS;薯块膨大期氮钾配施对甘薯13C分配的影响直接作用系数较大的是n、ΦPSⅡ和SPS。【结论】生长前期,氮钾配施处理通过提高n、/和SS促进光合产物在地上部积累,实现“建源”,而薯块膨大期主要提高n、ΦPSⅡ和SPS促进光合产物由地上部向地下部转运,兼顾“促流”和“扩库”,氮钾协同最终提高了甘薯产量。
甘薯;光合特性;叶绿素荧光;13C分配;酶活性
0 引言
【研究意义】源和库是作物形成经济产量的两个关键因素,大多数作物库源关系均有建立和平衡的过程,甘薯具有典型的库源关系,协调库源关系是甘薯高产的保障[1]。氮素是影响甘薯库源关系建立、发展的重要因素[2-4]。在一定范围内,增施氮肥可以提高甘薯的干物质生产能力和块根产量,而施氮量过高会导致地上部旺长、延迟结薯、不利于薯块的膨大[5-7]。近年来随氮肥用量不断投入,土壤中的氮素往往超过了甘薯生长所需[8],有研究表明土壤碱解氮含量超过44.14 mg·kg-1时,施氮就会破坏甘薯正常的库源关系,造成产量的下降[9],这成为单位化肥施用量不低而增产效益不高的主要原因之一。钾素是作物“库-源”器官之间碳水化合物转移与分配的“润滑剂”,适量的钾素供应能够调控甘薯T/R值,促进光合产物向块根的转移[10],可缓解因氮素过高而引起的库源失衡问题。因此通过氮钾配施的方式来调控库源关系成为一种有益途径[11-12]。【前人研究进展】光合作用是甘薯利用光能合成光合产物最重要过程,其中光系统Ⅱ(PSⅡ)是比较敏感的部位,PSⅡ反应中心的功能是限制光能利用和光合作用正常运行的关键环节,而叶片叶绿素荧光诱导动力学可以在非离体条件下方便快速反映PSⅡ反应中心、电子供体侧和受体侧的功能变化[13-14]。通过对光合特性和叶绿素荧光动力学的研究,使人们能够从更微观的层次了解甘薯光合系统的光能吸收、转换和利用效率[15]。同时,经光合作用同化的单糖由甘薯叶片中蔗糖合成酶(SS)和蔗糖磷酸合成酶(SPS)等碳代谢酶的催化形成蔗糖,光合产物以蔗糖的形式在甘薯植株内运输并积累。研究氮钾互作条件下叶片光合速率、叶绿素荧光参数和氮代谢酶活性的变化,对于揭示氮钾配施对甘薯光合产物分配积累的生理机制有重要的意义。利用同位素示踪技术研究植物对碳素的吸收与分配已相当成熟,在甘薯特定生长时期进行13CO2的定量标记,研究甘薯对碳素转移分配的速率及转移量,是一种瞬时效应的反映,能够清晰并定量反映碳素运转的过程。氮钾配施对大豆[16]、小麦[17]等作物生长发育、代谢过程和产量的影响已有广泛报道,而有关甘薯的氮钾配施研究相对较少。贾赵东等[18]和Tyler等[19]研究发现钾氮合理配施有利于库器官中干物质积累量,提高甘薯产量。汪顺义等[20]和江燕等[21]认为氮钾配施能显著提高甘薯氮和钾的养分积累量,提高甘薯源器官光合产物积累量。王萌等[22]认为氮钾配施能显著提高块根中蛋白质和可溶性糖类含量,改善甘薯品质。分析以上研究结果可以发现,甘薯的氮钾配施研究多集中于对养分吸收、产量和品质的调控效应,有关氮钾配施条件下甘薯不同生育期光合产物的积累分配过程研究相对较少。汪顺义等[23]对氮钾配施条件下氮素转移分配进行初探,发现施钾能够通过调控甘薯氮代谢酶来影响生长前期和薯块膨大期氮素在甘薯植株内的转移与分配。【本研究切入点】尽管调控氮素的分配可以实现库源关系的平衡,但对碳的调控才是库源平衡的本质,因此氮钾配施调控甘薯光合产物转移分配及其生理机制有待进一步探究。【拟解决的关键问题】本文以北方主栽甘薯品种商薯19号为材料,利用叶片13C标记技术,在甘薯生长前期和薯块膨大期,从光合特性和碳代谢酶活性角度探究氮钾配施对光合产物分配的影响及调控机制,为甘薯养分管理提供理论依据。
1 材料与方法
1.1 试验设计
试验选用北方主栽淀粉型甘薯品种商薯19号,于2014年5月15日和2015年5月17日在青岛农业大学胶州实验基地布置田间试验。试验区属于暖温带季风气候,降水集中,雨热同季,年平均气温11—14℃。土壤类型为砂姜黑土,试验开始前,采表层土壤(0—20 cm)风干磨碎后,用外加热重铬酸钾容量法测有机质,用NaOH扩散法测速效氮,NaHCO3浸提-钼锑抗比色法测有效磷,NH4OAc浸提-火焰光度法测速效钾。土壤养分含量见表1。

表1 不同试验年份移栽前土壤肥力状况
试验设置4个处理,分别为:CK(空白对照)、N75(专家推荐施肥量:75 kg N·hm-2)、K150(专家推荐施肥量:150 kg K2O·hm-2)、N75K150(纯氮75 kg N·hm-2+150 kg K2O·hm-2),试验采用起垄净作栽培方式,株距0.22 m、垄距0.8 m,小区面积48 m2(7.5 m×6.4 m),每个处理3次重复,随机区组排列。
田间试验移栽后40 d和100 d,进行13C叶片标记。方法如下:在晴朗天10:00—11:00,从每个小区选择生长一致具有代表性的植株3株,在其主茎第五片展开叶上标记13CO2。13CO2由Ba13CO3(13C 99%)和磷酸在反应器中反应生成,并用气球收集;标记前将欲标记叶用体积约为500 mL的聚氯乙烯透明塑料薄膜袋密封,用医用注射器注入50 mL13CO2(1%),在自然光照下光合同化一个小时之后撤掉塑料薄膜袋,标记完成3 d后将标记的3株植株收获。
1.2 测定项目与方法
干物质量:田间试验于40、75、100、120和150 d进行田间采样,将植株分为地上部和地下部,105℃杀青30 min,然后75℃烘干至恒重,并计算冠根比(地上部干物质量/地下部干物质量),于155 d进行收获,每小区选取200株甘薯进行统计并计算产量。
样品全碳含量及13C丰度测定:各部位用蒸馏水洗净后烘干磨碎,用同位素比率质谱仪(DELTA V Advantage)在中国农业科学院环发所稳定性同位素实验室测定13C及全碳含量(C,%)。计算方法如下:
Fi(13C丰度,%)=(δ13C + 1000)×R标/[(δ13C+1000)×R标+1000]×100,R标为碳同位素的标比值,R标= 0.0112372;
各器官13C积累量13Ci = Ci×(Fi-F自然)×10;
总碳积累量(g)=生物量(g)×甘薯器官全碳含量(%);
13C分配率(%)=该器官13C积累量/植株总13C积累量×100%。
净光合速率与SPAD值测定:于移栽后40 d和100 d上午9:00—10:30采用CIRAS-3便携式光合测定仪(Hansatech,USA),人工控制CO2浓度400 μmol CO2·mol-1、温度25℃、光照强度1 200 μmol·m-2·s-1,测定净光合速率(n)、胞间CO2浓度(i)、气孔导度(s)和蒸腾速率(r)。用SPAD仪测定第5片功能叶SPAD值。
荧光参数测定:SPAD值可表征甘薯功能叶总叶绿素相对含量。PSⅡ最大光化学效率(v/m)是反映PSⅡ活性中心的光能转换效率参数,PSⅡ实际光化学效率(ΦPSⅡ)表示电子在PSⅡ和PSⅠ间的传递情况,光化学淬灭系数(qP)通常用于反映PSⅡ原初电子受体QA的氧化还原状态和PSⅡ开放中心的数目,qP越大,说明PSⅡ的电子传递活性越大。ΦCO2表示CO2同化速率相对应的荧光量子产量,反映CO2同化速率。测定方法如下:叶片先进行15 min的暗适应处理,然后采用FMS-2便携式脉冲调制式荧光仪(英国Hansatech公司生产)测定叶片快速叶绿素荧光诱导动力学曲线(O-J-I-P曲线)。随后利用JIP-test对O-J-I-P曲线进行分析,解析0、v、m、s、m’和v’ 等叶绿素荧光参数。
ΦPSⅡ=(Fm’-Fs)/Fm’;
qP= (Fm’-Fs)/(Fm’-F0’)。
式中,ΦPSⅡ为PSⅡ实际光化学效率;qP为光化学淬灭系数。
SS和SPS酶活性测定:参照Doehlert[24]等的方法测定。称取0.5 g样品,加入7 mL Hepes-NaOH缓冲液(pH7.5),冰浴研磨至匀浆,匀浆转入15 mL离心管,10 000×4℃离心10 min,上清液即为酶液。取50 μL 酶液加入50 μL缓冲液、40 μL 25 mmol·L-1MgCl2、40 μL 50 mmol·L-1UDPG和20 μL100 mmol·L-16-磷酸果糖(果糖),30℃保温30 min,100℃沸水浴1 min,加入100 μL 2 mol·L-1NaOH混匀,100℃水浴10 min,加入2.0 mL 30%HCl和1 mL 1%间苯二酚,混匀后80℃保温10 min,冷却后480 nm波长下比色,测定生成磷酸蔗糖量,用蔗糖生成量表示酶活性。酶液经沸水浴后加入反应液为对照。
1.3 数据分析方法
为明确处理间效应差异,用SPSS18.0进行单因素方差分析分析,LSD法比较平均数间的差异显著性;为检验氮、钾两因素交互效应是否达到显著性水平,用DPS7.05软件双因素分析。甘薯光合产物的积累受光合特性、叶绿素荧光特性影响,光合产物分配主要由碳代谢酶活性调控,为明确各指标光合产物积累分配的直接作用大小,利用DPS7.05软件进行逐步回归分析与通径分析。
2 结果
2.1 氮钾配施对甘薯产量的影响
由表2所示,两年度试验,与N75和K150处理相比氮钾配施处理显著增加甘薯产量(<0.05),2014年增产幅度为10.1%和7.9%,2015年增产幅度为10.7%和9.3%。双因素分析表明,氮钾对甘薯产量的影响存在显著的正交互作用,其中,2014年交互效应值为0.95 t·hm-2,2015年交互效应值为1.35 t·hm-2(AB交互效应=(AB处理效应-A处理效应-B处理效应)/2)。两年田间试验甘薯产量结果相似,且年际间呈现递增趋势。

表2 氮钾配施对甘薯产量的影响
数据格式为平均值±误差。不同小写字母表示处理间差异显著(<0.05)。*,<0.05;**,<0.01;NS,>0.05
The data format is the average±deviation. Different small letters mean significant difference among treatments at 0.05 level. *,<0.05; **,<0.01; NS,>0.05
2.2 氮钾配施对甘薯冠根比(T/R)的影响
如表3所示,两年度试验,与单独施氮和单独施钾处理相比,氮钾配施处理提高了移栽后40 d和75 d冠根比,这表明氮钾互作提高了甘薯生长前期光合产物向地上部的分配;移栽100 d以后,氮钾配施处理冠根比均低于单独施氮和单独施钾处理,这表明氮钾互作有利于甘薯薯块膨大期由地上部向地下部转移。双因素分析表明,氮钾对冠根比存在显著的交互效应(<0.05),且各生育期互作效应明显(<0.05)。
2.3 氮钾配施对甘薯13C积累量的影响
方差分析表明2014和2015两年的年际间数据差异不显著,故用2015年数据表示(表4)。生长前期(40 d),与CK相比,施氮处理(N)和施钾处理(K)均能提高甘薯地上部和地下部13C分配量。与N、K处理相比,氮钾配施处理(N+K)地上部13C积累量分别提高42.3%和63.3%(<0.05);地下部13C积累量分别提高7.5%和18.5%。双因素分析可见,氮钾对地上部和地下部13C积累量存在显著的正交互效应(<0.05),其中,地上部13C积累量交互效应值为7.75 µg/plant,地下部13C积累量交互效应值为0.95 µg/plant。由13C分配率来看,氮钾配施处理显著提高了地上部13C分配率(达60.7%),有利于此时期13C在地上部的分配积累。薯块膨大期(100 d),施氮处理(N)和施钾处理(K)均能显著提高甘薯地上部和地下部13C分配量(<0.05),与N、K处理相比,氮钾配施处理(N+K)地上部13C积累量分别提高9.6%和18.7%;地下部13C积累量分别提高47.1%和38.8%(<0.05)。双因素分析表明,氮钾对地上部13C积累量交互效应不显著,但对地下部13C积累量存在显著的正交互效应,效应值为11.6 µg/plant。由13C分配率来看,氮钾配施处理降低了地上部13C分配率,增加了地下部13C分配率(达71.6%),促进了13C由地上部向地下部的转移。

表3 氮钾配施对甘薯冠根比值的影响
***,0.001水平显著 means significant difference at 0.001 level
2.4 氮钾配施对甘薯功能叶光合特性的影响
双因素分析表明,N、K处理均显著提高了甘薯两关键生长期净光合速率(n)、气孔导度(s)和胞间CO2浓度(i),且氮钾存在显著的交互作用(<0.05),但对蒸腾速率的影响交互效应不显著(表5)。生长前期(40 d),与N、K处理相比,氮钾配施处理n分别提高20.2%和19.6%,交互效应值为1.45 µmol CO2·m-2·s-1;s提高33.8%和20.0%;i分别提高13.1%和17.3%。薯块膨大期(100 d),与N、K处理相比,氮钾配施处理n分别提高9.1%和12.8%,交互效应值为0.20 µmol CO2·m-2·s-1;s提高33.6%和23.9%;i分别提高12.2%和17.0%。由变化幅度来看,氮钾交互作用主要调控了s变化从而显著增加n。

表4 氮钾配施对甘薯13C积累量的影响
2.5 氮钾互配施对甘薯叶绿素荧光特性的影响
生长前期(40 d),双因素分析表明氮、钾对SPAD、v/m、ΦPSⅡ、qP和ΦCO2存在显著正交互效应影响(<0.05)。与N、K处理相比,氮钾配施处理CO2同化速率(ΦCO2)分别提高30.4%和39.7%,其交互效应值为9.0 mmol·m-2·s-1。在薯块膨大期(100 d),双因素分析可见,N和K均显著提高了SPAD、Fv/Fm、ΦPSⅡ、qP和ΦCO2(<0.05),且氮钾存在显著的交互效应(<0.05)。与N、K处理相比,氮钾配施处理CO2同化速率(ΦCO2)分别提高27.1%和33.9%,其交互效应值为16.6 mmol·m-2·s-1(表6)。

表5 氮钾配施对甘薯叶片光合特性的影响

表6 氮钾配施对甘薯叶片叶绿素荧光特性影响
2.6 氮钾配施对甘薯SS和SPS酶活性的影响
甘薯光合产物的运输与分配由源-库器官间膨压决定,以磷酸蔗糖合成酶(SPS)和蔗糖合成酶(SS)为主的碳代谢酶活性通过调控蔗糖合成速率进而调控膨压差。由表7可见,双因素分析表明,施氮显著提高两生长期功能叶和块根SS和SPS酶活性(<0.05),施钾极显著提高了SS和SPS酶活性(<0.01),氮钾对两时期SS和SPS有显著的交互效应(<0.05)。在生长前期(40 d),相同处理条件下叶片中SS和SPS活性高于块根,而薯块膨大期(100 d)则呈相反趋势。这表明,在甘薯生长前期,氮钾配施处理通过提高叶片中SS和SPS酶活性,促进光合产物在代谢活性相对较高的源器官分配;而薯块膨大期,氮钾互作条件下块根中代谢活性显著高于叶片,光合产物在蔗糖浓度压力差的作用下由源器官向库器官运转。
2.7 逐步回归分析与通径分析
以甘薯SPAD值(1)、n(2)、v/m(3)、ΦPSⅡ(4)、qP(5)、SS(6)和SPS(7)为自变量,甘薯13C分配量(1:40 d地上部13C积累量;2:100 d地下部13C积累量)为因变量进行的逐步回归分析,其中,40 d回归关系为1=-9.894+0.0331- 0.0482-42.6563+30.1934+17.8335-0.1186+0.0127(1=0.954,=654.3,=0.021),100 d回归关系为2=-52.622-15.9081+6.6222+655.9583+1.3254- 0.8375+1.4276+0.2457(2=0.912,=578.2,= 0.032),说明以上指标是影响甘薯13C分配的主要因素。为进一步明确逐步回归确定的指标对13C分配差异的调控效应,本研究进行了通径分析(表8)。结果表明,生长前期氮钾配施对甘薯13C分配的影响直接作用系数最大的是n、v/m和SS;薯块膨大期氮钾配施对甘薯13C分配的影响直接作用系数最大的是n、ΦPSⅡ和SPS。这表明在生长前期n、Fv/Fm 和SS对甘薯13C分配的影响起主导作用,在薯块膨大期n、ΦPSⅡ和SPS起主导作用。

表7 氮钾配施对甘薯SS和SPS酶活性的影响
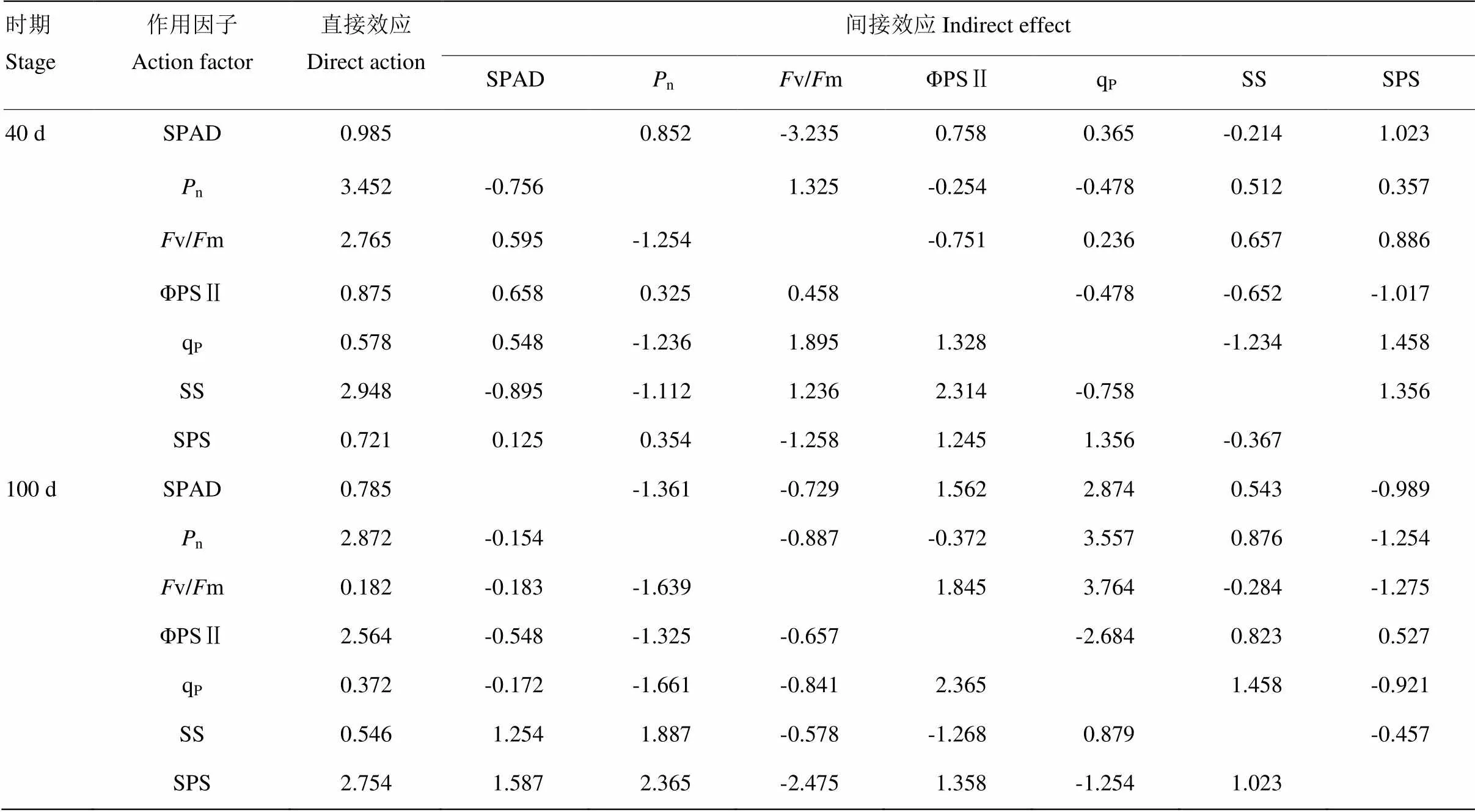
表8 氮钾配施作用对甘薯13C分配量与生理指标的通径系数
3 讨论
3.1 氮钾配施通过提高光合特性加速光合产物积累
叶绿素荧光动力学参数能准确反映甘薯叶片光能吸收的分配去向[25],其中光化学猝灭系数反映了光能向光合碳同化方向转移的比率,ΦCO2等参数能表征原初反应中CO2同化速率[26]。本试验结果表明,与单独施氮和单独施钾处理相比,氮钾配施处理显著提高了甘薯两生长时期v/m、ΦPSⅡ、qP和ΦCO2(<0.05)。这表明,一方面,氮钾配施处理能显著增加源于光合磷酸化和电子传递过程相偶联所生成的ATP和NADPH的量,进而促进碳同化过程[27]。另一方面,充足供应氮钾能够使叶片较大部分反应中心保持开放状态[28],QA处于较高的还原态,这有利于快速有效的传递H2O光解产生的电子[29],氮钾配施条件下PSⅡ过程得到显著的促进。PSⅡ反应中心功能的升高,能显著提高光合机构吸收的光能进入光化学过程的量,提高了光能向碳同化转移的比率[30]。叶绿素荧光是研究甘薯光合作用敏感的探针,光合碳同化速率的提高必然会引起光合作用的变化。
从氮钾配施对光合特性的影响来看,一方面,氮钾配施处理条件下充足的氮素有助于甘薯叶片叶绿素的合成,充足的钾素利于气孔导度的提高[31],减小了CO2从细胞间隙向叶绿体传递阻力并使碳同化过程CO2的利用增加(包括碳同化酶活性和能力合成等过程);另一方面,氮钾配施处理能显著提高Rubisco酶活性[32],最终提高甘薯两关键生长时期光合速率[33]。氮钾配施处理通过提高PSⅡ和n等光合途径增加光合产物的同化量。本试验干物质积累量的研究结果表明,氮钾配施处理生长前期地上部13C积累量增加42.3%—63.3%(<0.05),薯块膨大期地上部13C积累量增加9.6%—18.7% (<0.05)。
3.2 氮钾配施通过影响碳代谢酶活性调控光合产物的分配
光合作用积累的光合产物(13C)在甘薯植株内的运输与分配主要由源-库器官间膨压决定[34],以SPS和SS为主的碳代谢酶活性通过调控蔗糖合成速率进而调控膨压差[35]。本试验发现,氮钾对两时期内SS和SPS存在显著的正交互作用(<0.05)。在甘薯生长前期,氮钾配施条件下叶片中SS和SPS酶活性显著高于块根,13C多分配于代谢活性旺盛的源器官,并显著提高了移栽75 d之前冠根比;而薯块膨大期,氮钾配施处理通过提高块根中SS和SPS酶活性,加速块根中蔗糖积累速率,13C在库-源器官膨压差的作用下由叶片向块根中分配,促进了块根中光合产物积累量,显著降低了移栽100 d之后冠根比。
3.3 甘薯氮钾互作效应模型参数分析
对氮钾配施条件下光合产物积累与分配的作用因子的贡献度进行通径分析,生长前期氮钾配施对甘薯13C分配的影响直接作用系数最大的是n、v/m和SS,而薯块膨大期甘薯13C分配的影响直接作用系数最大的是n、ΦPSⅡ和SPS。这表明对甘薯13C分配的影响在生长前期n、v/m 和SS起主导作用,促进13C在地上部积累;在薯块膨大期n、ΦPSⅡ和SPS起主导作用,促进13C由地上部向地下部转运。在氮钾互作条件下如何调控光合产物在植株内的转移运输过程,则须结合离子通道和生理解剖结果进行进一步探讨。
3.4 氮钾配施通过调控甘薯光合产物积累与分配进而协调库源关系
氮钾互作条件下不同生长期内光合产物转移和分配的影响存在差异。本研究表明,氮钾配施处理在生长前期(40 d)能显著提高13C在地上部积累量(<0.05),13C分配率也显著高于单独施氮和单独施钾处理,高达60.7%。而薯块膨大期(100 d)与生长前期(40 d)变化趋势相反,氮钾配施处理显著提高了R/T值,13C地下部分配率高达71.6%(<0.05)。甘薯生长前期是库源关系建立初期,地上部需不断增加源规模以满足生长后期块根膨大所需,此时代谢中心是甘薯的地上部(叶片)[36]。氮钾配施处理促进生长前期光合产物向地上部“源”器官分配,为甘薯地上部提碳素骨架以保证甘薯植株形态的构建。到了薯块膨大期,地上部光合产物开始向块根转移,此时块根成为新的代谢中心[37]。氮钾配施处理促进薯块膨大期氮素向地下部“库”转移,即实现了“促流”同时又达到了“扩库”目的[38]。氮钾互作效应调控光合产物在关键生长期内合理分配进而协调了甘薯库源关系。
4 结论
氮钾平衡施肥条件下,氮通过提高光合活性增加光合产物积累,钾因素通过提高碳代谢酶活性加速光合产物的运输。生长前期,氮钾配施处理通过提高n、v/m和SS促进光合产物在地上部积累,实现“建源”,而薯块膨大期主要提高n、ΦPSⅡ和SPS促进光合产物由地上部向地下部转运,兼顾“促流”和“扩库”,氮钾配施能协同光合产物的积累与分配,最终提高甘薯产量。
References
[1] Haimeirong, Kubota F. The effects of drought stress and leaf ageing on leaf photosynthesis and electron transport in photosystem 2 in sweet potato (Lam.) cultivars., 2003, 41(2): 253-258.
[2] 史春余, 张晓冬, 张超, 陈晓光. 甘薯对不同形态氮素的吸收与利用. 植物营养与肥料学报, 2010, 16(2): 389-394.
SHI C Y, ZHANG X D, ZHANG C, CHEN X G. Absorption and utilization of different nitrogen forms for sweet potato., 2010, 16(2): 389-394. (in Chinese)
[3] 宁运旺, 曹炳阁, 马洪波, 汪吉东, 张辉, 许仙菊, 张永春. 氮肥用量对滨海滩涂区甘薯干物质积累、氮素效率和钾钠吸收的影响. 中国生态农业学报, 2012, 20(8):982-987.
NING Y W, CAO B G, MA H B, WANG J D, ZHANG H, XU X J, ZHANG Y C. Effects of nitrogen application rate on dry matter accumulation, nitrogen efficiency, and potassium and sodium uptake of sweet potato () in coastal North Jiangsu province., 2012, 20(8): 982-987. (in Chinese)
[4] 唐忠厚, 李洪民, 张爱君, 史新敏, 魏猛, 陈晓光, 丁艳锋. 甘薯叶光合特性与块根主要性状对氮素供应形态的响应. 植物营养与肥料学报, 2013, 19(6): 1494-1501.
TANG Z H, LI H M, ZHANG A J, SHI X M, WEI M, CHEN X G, DING Y FResponses of nitrogen supply forms on leaf photosynthetic characteristics and root characters of sweet potato., 2013, 19(6): 1494-1501. (in Chinese)
[5] OSAKI M, UEDA H, SHINANO T, MATSUI H, TADANO T. Accumulation of carbon and nitrogen compounds in sweet potato plants grown under deficiency of N, P, or K nutrients., 1995, 41(3):557-566.
[6] Hartemink A E. Integrated nutrient management research with sweet potato in Papua New Guinea.2003, 32(3): 173-182.
[7] Ankumah R O, Khan V, Mwamba K, Kpomblekou-A KThe influence of source and timing of nitrogen fertilizers on yield and nitrogen use efficiency of four sweet potato cultivars., 2003, 100(2): 201-207.
[8] 周全卢. 秋甘薯不同类型品种干物质积累特性研究[D]. 重庆: 西南大学, 2007.
ZHOU Q L. Research of dry matter accumulating characters on autumn sweet potato varieties of different types[D]. Chongqing: Southwest University, 2007.
[9] 高璐阳, 房增国, 史衍玺. 施氮量对鲜食型甘薯产量、品质及氮素利用的影响. 华北农学报, 2014, 29(6):189-194.
GAO L Y, FANG Z G, SHI Y X. Effects of N application rate on yield, quality and the influence of nitrogen utilization of sweet potato., 2014, 29(6): 189-194. (in Chinese)
[10] 宁运旺, 曹炳阁, 朱绿丹, 张永春, 汪吉东, 许仙菊, 张辉, 马洪波. 施钾水平对甘薯干物质积累与分配和钾效率的影响. 江苏农业学报, 2012, 28(2): 320-325.
NING Y W, CAO B G, ZHU L D, ZHANG Y C, WANG J D, XU X J, ZHANG H, MA H B. Effects of potassium application rates on dry matter accumulation, dry matter distribution, and potassium efficiency of sweet potato., 2012, 28(2): 320-325. (in Chinese)
[11] 史春余, 王振林, 赵秉强,郭风法, 余松烈. 钾营养对甘薯块根薄壁细胞微结构、14C同化物分配和产量的影响. 植物营养与肥料学报, 2002, 8(3): 335-339.
SHI C Y, WANG Z L, ZHAO B Q,GUO F F, YU S LEffects of potassium on the parenchyma cell structure of storage root, the distribution of14C photo synthates and yield in sweet potato., 2002, 8(3): 335-339. (in Chinese)
[12] 李绍长, 王荣栋. 作物源库理论在产量形成中的应用. 新疆农业科学, 1998, 6(3): 106-110.
LI S C, WANG R D. Application of crop source sink theory in yield formation., 1998, 6(3): 106-110. (in Chinese)
[13] Shcansker G, Srivastava A, Covindjee, Strasser R J. Characterization of the 820-nm transmission signal paralleling the chlorophyll a fluorescence rise (OJIP) in pea leaves., 2003, 30(7): 785-796.
[14] Strasser R J, Srivastava A, Covindjee. Polyphasic chlorophyll a fluorescence transients in plants and cyanobacteria.1995, 61(1): 32-42.
[15] 李鹏民, 高辉远, Strasser R J.快速叶绿素荧光诱导动力学分析在光合作用研究中的应用.植物生理与分子生物学学报, 2005, 31(6): 559-566.
LI P M, GAO H Y, Strasser R J. Application of the fast chlorophyll fluorescence induction dynamics analysis in photosynthesis study.2005, 31(6): 559-566. (in Chinese)
[16] 邹铁祥, 戴廷波, 姜东,荆奇, 曹卫星. 钾素水平对小麦氮素积累和运转及籽粒蛋白质形成的影响. 中国农业科学, 2006, 39(4): 686-692.
ZOU T X, DAI T B, JIANG D, JING Q, CAO W X. Potassium supply affected plant nitrogen accumulation and translocation and grain protein formation in winter wheat., 2006, 39(4): 686-692. (in Chinese)
[17] 汪自强, 陶秀明, 董明远. 不同供钾水平下春大豆的氮积累和利用. 中国农业科学, 1997, 30(5): 20-25.
WANG Z Q, TAO X M, DONG M Y. Nitrogen accumulation and utilization of various spring soybean genotypes under different potassium supplies., 1997,30(5): 20-25. (in Chinese)
[18] 贾赵东, 马佩勇, 边小峰,郭小丁, 谢一芝. 氮钾配施和栽插密度对甘薯干物质积累及产量形成的影响. 华北农学报, 2012, 27(S1): 320-327.
JIA Z D, MA P Y, BIAN X F,GUO X D, XIE Y Z. Effects of different N and K fertilizer ratio and planting density on yield and dry matter accumulation of sweet potato., 2012, 27(S1): 320-327. (in Chinese)
[19] Monday T A, Foshee III W G, Blythe E K, DOZIER J R, WELLS L W, Sibley J L, BROWN J ENitrogen requirements for sweet potato following a crimson clover cover crop., 2013, 19(1): 78-82.
[20] 汪顺义, 李欢, 刘庆, 史衍玺. 氮钾互作对甘薯根系发育及碳氮代谢酶活性的影响. 华北农学报, 2015, 30(5): 167-173.
WANG S Y, LI H, LIU Q, SHI Y XInteractive effects of nitrogen and potassium on root growth and leaf enzyme activities of sweet potato., 2015, 30(5): 167-173. (in Chinese)
[21] 江燕. 氮钾互作对甘薯养分吸收利用和产量的影响[D]. 泰安: 山东农业大学, 2015.
JIANG Y.Effect of nitrogen and potassium interaction on nutrient absorption, utilization, and yield of sweet potato[D]. Tai’an: Shandong Agriculture University, 2015. (in Chinese)
[22] 王萌, 房增国, 梁斌, 曾路生, 李俊良. 高肥力土壤氮钾配施对鲜食型甘薯产量及品质的影响. 华北农学报, 2016, 31(5): 199-204.
WANG M, FANG Z G, LIANG B, ZENG L S, LI J L. Effects of combined application of nitrogen and potassium fertilizer on yield and quality of fresh-eating sweet potato in soil with high fertility., 2016, 31(5): 199-204. (in Chinese)
[23] 汪顺义, 刘庆, 史衍玺, 李欢. 施钾对甘薯氮素转移分配及氮代谢酶活性的影响. 应用生态学报, 2016, 27(11): 3569-3576.
WANG S Y, LIU Q, SHI Y X,LI H. Effects of potassium on nitrogen translocation and distribution and nitrogen metabolism enzyme activities of sweet potato., 2016, 27(11): 3569-3576. (in Chinese)
[24] Doehlert D C, Kuo T M, Felker F C. Enzymes of sucrose and hexose metabolism in developing kernels of two inbreds of maize.,1988, 86(4): 1013-1019.
[25] Li P M, Cai R G, Gao H Y, Peng T, Wang Z LPartitioning of excitation energy in two wheat cultivars with different grain protein contents grown under three nitrogen applications in the field., 2007, 129(4): 822-829.
[26] 张善平, 冯海娟, 马存金,李耕, 刘鹏, 董树亭, 赵斌, 张吉旺, 杨今胜. 光质对玉米叶片光合及光系统性能的影响. 中国农业科学, 2014, 47(20): 3973-3981.
ZHANG S P, FENG H J, MA C J, LI G, LIU P, DONG S T, ZHAO B, ZHANG J W, YANG J S. Effect of light quality on photosynthesis and photosystem of maize (L.) leaves., 2014, 47(20): 3973-3981. (in Chinese)
[27] Gilmore A M, Hazlett T L, Debrunner P G. Comparative time-resolved photosystem II chlorophyII a fluorescence analyses reveal distinctive differences between photoinhibitory reaction center damage and xanthophyll cycle-dependent energy dissipation., 1996, 64(3): 552-563.
[28] Yu G S, Liu B, Lan F W, Li M H, LIU Y. Damage to the oxygen-evolving complex by superoxide anion, hydrogen peroxide, and hydroxyl radical in photoinhibition of photosystem II., 2006, 90(1): 67-78.
[29] 孙骏威, 李素芳, 付贤树, 奚辉, 王腾浩. 低钾对水稻不同叶位叶片光合特性及抗氧化系统的影响. 核农学报, 2006, 21(4): 404-408.
SUN J W, LI S F, FU X S, XI H, WANG T H. Effects of low potassium stress on photosynthetic characteristics and antioxidant systems in different position leaves of rice plants., 2006, 21(4): 404-408. (in Chinese)
[30] Bednarz C W, Oosterhuis D M, Evans R D. Leaf photosynthesis and carbon isotope discrimination of cotton in response to potassium deficiency., 1998, 39(2): 131-139.
[31] BATTIE-LACLAU P, LACLAU J P, BERI C, CONSTANCE BERI, LAURIANE MARTA R, ALMEIDA MUNIZ, BRUNA CERSOZIMO ARENQUE, MARISA DE CASSIA PICCOLO, LIONEL JORDAN- MEILLE, JEAN-PIERRE BOUILLET, YANN NOUVELLON.Photosynthetic and anatomical responses of eucalyptus grandis leaves to potassium and sodium supply in a field experiment., 2014, 37(1): 70-81.
[32] 郑炳松, 程晓建, 蒋德安, 翁晓燕. 钾元素对光合速率、Rubisco和RCA的影响. 浙江林学院学报, 2002, 19(1): 104-108.
ZHENG B S, CHENG X J, JIANG D A, WENG X Y. Effects of potassium on Rubisco, RCA and photosynthetic rate of plant.2002, 19(1): 104-108. (in Chinese)
[33] 孙骏威, 翁晓燕, 李峤, 邵建林. 缺钾对水稻不同品种光合和能量耗散的影响. 植物营养与肥料学报, 2007, 13(4): 577-584.
SUN J W, WEN X Y, LI Q, SHAO J L. Effects of potassium- deficiency on photosynthesis and energy dissipation in different rice cultivars., 2007, 13(4): 577-584. (in Chinese)
[34] 柳洪鹃, 姚海兰, 史春余, 张立明. 施钾时期对甘薯济徐23块根淀粉积累与品质的影响及酶学生理机制. 中国农业科学, 2014, 47(1): 43-52.
LIU H J, YAO H L, SHI C Y, ZHANG L M. Effect of potassium application time on starch accumulation and related enzyme activities of sweet potato variety Jixu 23., 2014, 47(1): 43-52. (in Chinese)
[35] 崔光军, 刘风珍, 万勇善. 花生荚果干物质积累与蔗糖代谢的相关性研究. 中国农业科学, 2010, 43(19): 3965-3973.
CUI G J, LIU F Z, WAN Y S. Relationship between dry matter accumulation and sucrose metabolism during pod development in peanut(L.)., 2010, 43(19): 3965-3973. (in Chinese)
[36] 宁运旺, 马洪波, 张辉, 汪吉东, 许仙菊, 张永春. 甘薯源库关系建立、发展和平衡对氮肥用量的响应. 作物学报, 2015, 41(3): 432-439.
NING Y W, MA H B, ZHANG H, ZHANG J D, XU X J, ZHANG Y C.Response of sweetpotato in source-sink relationship establishment, expanding, and balance to nitrogen application rates., 2015, 41(3): 432-439. (in Chinese)
[37] 史春余, 王振林, 赵秉强, 郭风法, 余松烈. 钾营养对甘薯某些生理特性和产量形成的影响. 植物营养与肥料学报, 2002, 8(1): 81-85.
SHI C Y, WANG Z L, ZHAO B Q, GUO F F, YU S L. Effect of potassium nutrition on some physiological characteristics and yield formation of sweet potato., 2002, 8(1): 81-85. (in Chinese)
[38] 柳洪鹃, 史春余, 张立明, 张海峰, 王振振, 柴沙沙. 钾素对食用型甘薯糖代谢相关酶活性的影响. 植物营养与肥料学报, 2012, 18(3): 724-732.
LIU H J, SHI C Y, ZHANG L M, ZHANG H F, YU Z Z, CHAI S S. Effect of potassium on related enzyme activities in sugar metabolism of edible sweet potato., 2012, 18(3): 724-732. (in Chinese)
(责任编辑 杨鑫浩)
Interactive Effects of Nitrogen and Potassium onPhotosynthesis Product Distribution and Accumulation of Sweet Potato
WANG ShunYi, LIU Qing, SHI YanXi, LI Huan
(College of Resources and Environmental Science, Qingdao Agricultural University, Qingdao 266109, Shandong)
【Objective】The objective of this paper is to explore the interactive effects of nitrogen and potassium on photosynthesis product transfer allocation and its physiological mechanism of sweet potato.【Method】The two years field experiments include four treatments: CK, N, K and N+K,13C blade tag on leaves was made on 40 d and 100 d after transplanting. The interactive effects of nitrogen and potassium on13C distribution, photosyntheticcharacteristics, fluorescencecharacteristic, SS activities, SPS activities, and yield of sweet potato were studied. 【Result】Compared with N, K, interactions of N and K treatment, in 2014, yield increased by 10.1% and 7.9%, and the interactive effect value was 0.95 t·hm-2; in 2015, yield increased by 10.7% and 9.3%, and the interactive effect value was 1.35t·hm-2. Compared with N and K treatments, interactions of N and K treatment improved the functional leaf photosynthesis and chlorophyll fluorescence characteristics, and the accumulation of photosynthetic products increased at two key stages of sweet potato. For example, ΦCO2increased by 27.1%-39.7%,nincreased by 9.1%-20.2%, and13C total accumulation increased by 26.3%-42.2%. At the early stage of growth, nitrogen and potassium interactions treatment significantly increased leaf SS and SPS enzyme activities, their activities were significantly higher than that in root, and significantly improved the13C distribution rate in leaves by 60.7%, promoted photosynthetic products distribution in the source organ.In root rapid expansion period, interactions of N+K treatment significantly increased the SS and SPS enzyme activities of root, which were significantly higher than that of leaf blade, photosynthetic product distributed from the ground to the underground part under the action of pressure in the sink - source organs, nitrogen and potassium interactions increased the13C distribution rate in root by 71.6%. Stepwise regression analysis showed that the carbon metabolism enzyme photosynthetic characteristics and chlorophyll fluorescence characteristics are the key indicators of product distribution of sweet potato(1=0.954,2=0.912). Path analysis indicated that the largest coefficients affected13C distribution under nitrogen and potassium interaction weren,/and SS at the early stage of growth while key coefficients affected13C distribution weren, ΦPSⅡ and SPS in root rapid expansion period. 【Conclusion】It was concluded that nitrogen and potassium interaction promoted the accumulation of photosynthetic products in aerial part through increase ofn, ΦPSⅡ and SS, thus realized the construction of source at the early stage of growth. In root rapid expansion period, N and K interaction mainly improvedn, ΦPS and SPS and thus promoted the transfer of photosynthetic products from the ground to the underground part and promoted the yield of sweet potato.
sweet potato; photosynthetic characteristics;chlorophyll fluorescence;13C distribution; enzyme activities
2016-11-04;接受日期:2016-12-23
国家现代农业产业技术体系建设专项(CARS-11-B-14)、国家自然科学青年基金(41501271)
汪顺义,E-mail:644393504@qq.com。通信作者李欢,E-mail:lihuancomcomcom@163.com
猜你喜欢
作物学报(2022年8期)2022-05-29
湖北农机化(2021年11期)2021-12-07
保鲜与加工(2021年1期)2021-02-06
今日农业(2019年16期)2019-01-03
中成药(2018年2期)2018-05-09
现代园艺(2017年22期)2018-01-19
现代园艺(2017年19期)2018-01-19
农村百事通(2018年21期)2018-01-03
中国农业文摘-农业工程(2017年2期)2017-03-31
今日农业(2017年7期)2017-02-02
