
淹水胁迫对湖北海棠生长及光合作用的影响1)
2017-12-27 01:12张虎姜文龙武启飞张往祥曹福亮
东北林业大学学报 2017年12期
张虎 姜文龙 武启飞 张往祥 曹福亮
(江苏农林职业技术学院,句容,212400) (南京林业大学)
淹水胁迫对湖北海棠生长及光合作用的影响1)
张虎 姜文龙 武启飞 张往祥 曹福亮
(江苏农林职业技术学院,句容,212400) (南京林业大学)
以1年生湖北海棠实生苗为试验材料,研究了淹水胁迫对湖北海棠生长以及光合作用的影响。结果表明:在淹水初期(0~15 d),湖北海棠苗高生长受到的抑制作用不显著,而在淹水后期(15~35 d),抑制程度显著加剧,最终苗高表现为对照的10.7%;淹水胁迫整体对湖北海棠地径的抑制作用不显著。净光合速率、气孔导度以及气孔限制值在淹水期间均显著低于常规管理,而细胞间隙CO2摩尔分数受淹水的影响不显著;在淹水胁迫下,湖北海棠叶片花青素、类胡萝卜素相对质量分数显著高于对照,而叶绿素相对质量分数则显著低于对照。综上,湖北海棠是一种耐水性较强的苗木。
湖北海棠;淹水胁迫;光合作用
湖北海棠(Malushupehensis)为蔷薇科苹果属小乔木,是一种以观花、观果为主的观赏性树种[1],是中国不可多得的苹果属多抗性植物资源之一[2],具有较强的抗性[3],且易繁殖,嫁接成活率高,因而在苗木生产上常用作苹果砧木[4]。目前关于湖北海棠抗性的研究在抗寒性、抗病性等方面[5-6]较多,对于其耐水性方面的研究却少有报道,而涝害是我国较为常见的自然灾害之一,因此研究其耐水性将会对我国观赏海棠的生产,新品种开发与扩繁等具有重要意义。
植物光合作用是其生长及其他代谢活动的生理基础,不同的光合特性往往是植物生长、抗性等产生差异的直接原因之一[7]。淹水是植物较为常见的胁迫方式,多发生于江河流域、滩涂等多个生态系统[8],其对植物的生理学影响是多方面的[10]。因此加强对耐淹性树种的研究,培育具有强耐淹能力的树种,对洪涝灾害的防治意义重大。目前国内外对于淹水胁迫的研究主要包括:淹水胁迫的响应机制[10-13];淹水胁迫对植物生理生化的影响[14-15]以及淹水胁迫对植物光合作用的影响[16-19]。在湖北海棠的淹水胁迫研究方面多侧重于基因分子[20],鲜有关于淹水胁迫对湖北海棠光合作用影响的报道。
本研究通过分析湖北海棠在淹水胁迫下的光合生理指标及叶片色素指标的动态变化,研究了湖北海棠在淹水胁迫下生长、气体交换参数与叶片色素的动态变化规律,探讨了淹水胁迫对湖北海棠生长以及光合作用的影响,旨在为观赏海棠优异品种生产与开发、新品种扩繁及有效引种等提供理论依据与技术参考。
1 材料与方法
试验地位于江苏省扬州市江都区仙女镇。属于北亚热带季风气候,年均降水量1 000 mm,年均温14.9 ℃,无霜期约为320 d,地势平坦。试验地土壤质地为沙壤土,深厚肥沃。试验材料为湖北海棠一年生实生苗。2016年2月,选取长势相近的苗株300株,进行盆栽(口径30 cm,高30 cm),每盆3株,正常水分管理,培养6个月,至植株恢复正常稳定状态。
试验设对照组A(常规管理)和处理组B(淹水胁迫)两组。适应性栽培完成后,选择长势一致的种苗60株,分成两组,对照A组和处理B组各30株。将处理B组盆栽植株移入人工设置的水槽内进行淹水处理,并保持水位在土表以上5 cm,为防止水质恶化每3 d换水一次。分别于处理后0、5、10、15、20、25、30、35 d测定试验数据。对照A以及处理B均采用相同的测量方法。苗高和地径分别采用卷尺和游标卡尺进行测量,并计算其相对生长率。
下面对某220 kV GIS断路器合闸时间、合闸不同期时间严重超标、合闸速度偏低缺陷案例进行分析,排查、核实测试数据异常原因,并总结类似断路器操作机构检修经验,为提升断路器安装、验收、运维质量提供参考。
车速平稳连续降速依靠行车制动器无法完成,必须要借助缓速装置[8-9],它的工作原理与传统制动方式有所不同,它是在不使用或少使用行车制动器的情况下,减缓车辆行驶速度。目前,常用的持续制动装置主要有以下2种。
光合色素相对质量分数动态变化测定:采集5枚健康的湖北海棠叶片使用单通道光谱分析仪(UniSpec-SC)对叶片中色素组分的反射光谱值进行测定。每个叶片均匀选择6个不同的点进行测定,共重复30次。各色素相对质量分数按对应的光谱反射指标(Rx)计算:花青素(Anth)为R800(1/R550-1/R700),类胡萝卜素为R800(1/R520-1/R700),叶绿素为R705=(R750-R445)/(R705-R445)[21-22]。比例关系计算方法=W(叶绿素)/W(花青素+类胡萝卜素)。
光合特性动态变化测定:在2016年9月2日至10月11日,选择晴朗天气,在08:00—11:00期间采用光合仪(Li6400型)对湖北海棠进行光合指标的测定。所测叶片皆为生长健康的植株上部向阳面第3~5节位完全功能叶,每个品种重复3次,取平均值。测定参数包括:净光合速率(Pn)、气孔导度(Gs)、细胞间隙CO2摩尔分数(Ci),大气CO2摩尔分数(Ca)并根据测定的指标值计算气孔限制值(Ls)。气孔限制值(Ls)=(1-Ci)/(Ca)。
近年来,云南电网深入贯彻落实创新驱动发展战略,企业创新能力明显增强,积极开展了科技成果转化机制的探索与实践,公司在省部级科技进步奖励的层次和数量都在不断增加,科技创新、职工技术创新、QC活动等方面涌现出了一大批好的成果。
2 结果与分析
2.1 淹水胁迫对湖北海棠生长的影响
由表1结合图1可以看出,在苗高方面,湖北海棠在淹水后的5~15 d时苗高生长受抑制作用不显著,此时处理B与对照A的外部形态差别甚微。在淹水的20~35 d时,处理B的相对生长率开始上升,且处理B的叶片开始呈现出枯萎发黄态势,至后期部分植株叶片落光。苗高在此时受抑制作用虽较为显著,但与对照A相比差别不大。在整个淹水周期中,处理B与对照A的苗高相对增长率分别为10.70%和16.6%。在地径方面,处理B与对照A相比略有下降,处理B在淹水的第30 d受抑制作用显著,而其余时间受抑制不明显。在整个淹水周期中,处理B与对照A的地径相对增长率分别为7.91%和14.05%。
易地扶贫搬迁经验总结与思路——基于易地扶贫搬迁实地调查 …………………………………………………… 吕南辉(1/20)
表1湖北海棠在淹水胁迫下苗高和地径生长进程及相对生长率的变化
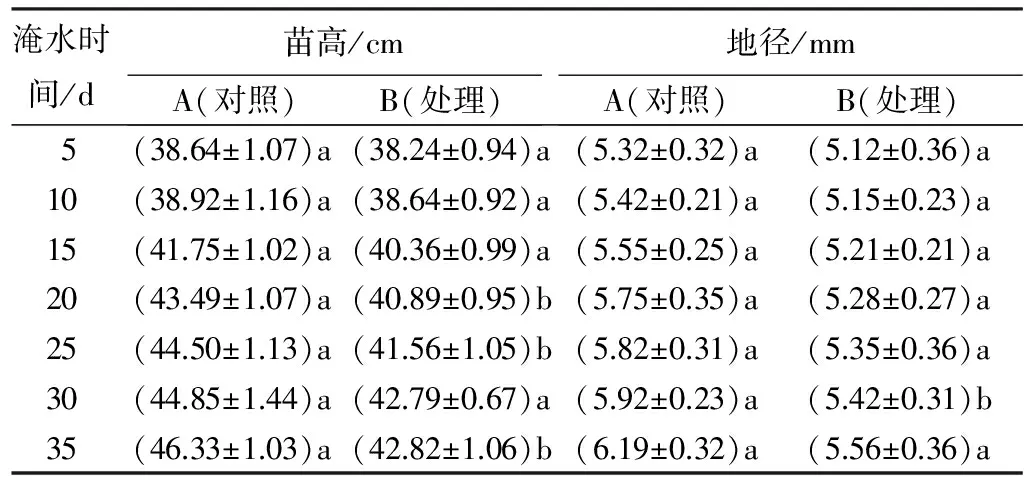
淹水时间/d苗高/cmA(对照)B(处理)地径/mmA(对照)B(处理)5(38.64±1.07)a(38.24±0.94)a(5.32±0.32)a(5.12±0.36)a10(38.92±1.16)a(38.64±0.92)a(5.42±0.21)a(5.15±0.23)a15(41.75±1.02)a(40.36±0.99)a(5.55±0.25)a(5.21±0.21)a20(43.49±1.07)a(40.89±0.95)b(5.75±0.35)a(5.28±0.27)a25(44.50±1.13)a(41.56±1.05)b(5.82±0.31)a(5.35±0.36)a30(44.85±1.44)a(42.79±0.67)a(5.92±0.23)a(5.42±0.31)b35(46.33±1.03)a(42.82±1.06)b(6.19±0.32)a(5.56±0.36)a
注:表中数据为平均值±标准差;同列不同字母表示差异显著(P<0.05)。
综上所述,从外部形态上看,湖北海棠在淹水的前期与对照A差别甚小,后期叶片有明显的枯萎发黄迹象,而苗高在淹水胁迫下后期较前期相比受抑制虽较为显著,但与对照A相差不大,总体上不受淹水影响;地径虽在个别天受抑制,但其余天数受抑制作用不显著,基本上不受淹水影响。
2.2 淹水胁迫对湖北海棠光合生理指标的影响
淹水胁迫下,气孔导度的的变化趋势基本和净光合速率相似,其指标显著低于对照,但在不同时期变化规律又表现出一定的差异性(表2)。淹水5、10 d时,处理B的气孔导度分别占对照A的47%和49%,而淹水15 d时其气孔导度占对照的80%,淹水25 d时气孔导度上升到整个处理过程的最大值(117 μmol·m-2·s-1),其孔导度占对照的90%。淹水30、35 d时气孔导度又开始下降,其孔导度分别占对照的50%和46%。
在淹水胁迫下,湖北海棠的净光合速率总体上呈先上升后下降的趋势。淹水5 d时,处理B在淹水胁迫下的净光合速率较对照A相比显著下降,其净光合速率占对照的21%。淹水10 d时,处理B在淹水胁迫下的净光合速率虽有所上升,但上升的幅度较小,其净光合速率占对照的39%。淹水15 d时,处理B的净光合速率继续上升,且上升到整个处理过程中的最大(5.34 μmol·m-2·s-1),其净光合速率占对照的54%。淹水20 d时,处理B的净光合速率再次下降,其净光合速率占对照的52%。淹水25、30、35 d时,处理B的净光合速率均有所下降,其净光合速率分别占对照的36%、39%和37%。

图1 湖北海棠形态表观变化
由表2可知,淹水胁迫下细胞间隙CO2摩尔分数与净光合速率表现出相反的变化规律,处理B在淹水的5、20 d时细胞间隙CO2摩尔分数显著高于对照A,其余时间不显著。淹水5 d时,处理B的细胞间隙CO2摩尔分数占对照的122%。淹水10 d时,处理B的细胞间隙CO2摩尔分数下降至整个处理过程的最小值(276.9 μmol·mol-1),且占对照的107%。淹水30 d时,处理B的细胞间隙CO2摩尔分数达到了最大值(327 μmol·mol-1),处理B的细胞间隙CO2摩尔分数占对照的107%。淹水35 d时,处理B的细胞间隙CO2摩尔分数和对照的基本相似。
在淹水胁迫下,花青素(P=0.001~0.092)、类胡萝卜素(P=0.001~0.291)的质量分数显著高于对照A,而叶绿素质量分数则低于对照A(P=0.043~0.172)。由表3可知,在淹水初期,叶绿素质量分数持续下降,而花青素和类胡萝卜素质量分数在5、10 d虽有下降,但总体呈上升趋势,20 d后花青素、类胡萝卜素和叶绿素的质量分数变化较初期相比趋于平稳。从色素占比关系图中可以看出,淹水后叶绿素的质量分数小于花青素与类胡萝卜素质量分数之和。
植物的生长指标(苗高、地径等)能够较好的判断出苗木受逆境影响的程度,这是评价苗木抗逆能力的最直接,最广泛的方法,而植物在逆境中的生长常会伴随一系列的形态结构变化[19]。

表2 湖北海棠净光合速率、气孔导度、细胞间隙CO2摩尔分数及气孔限制值动态变化
注:表中数据为平均值±标准差。
2.3 淹水胁迫对湖北海棠叶绿素质量分数的影响
基于花青素、类胡萝卜素、叶绿素的质量分数数据绘制了折线图,发现随着时间的推移,3大色素的相对含量及其比例关系呈现不同规律的变化。
综上所述,在应试教育当头的今天,我们是不是应当反思当前出现的教育问题.推动课程改革和教育改革的目的也是为了对抗应试教育,让更多的学生体会到学习本身的乐趣,让素质教育能够更快的替代应试教育.高中物理是高中学科中较为复杂的学科,教师在进行授课时不断对课堂教学进行改革,引导学生对学科进行探究,能够更好的推动物理学科在我国的发展.
气孔限制值Ls在一定程度上反映了植物叶片对大气CO2相对利用效率的大小,它与细胞间隙CO2摩尔分数密切相关,细胞间隙CO2摩尔分数越大,植物对大气的CO2利用率越大,气孔限制值就越小。由表2可以看出,处理B的气孔限制值与对照A相比总体上差异较为显著,其变化规律呈先上升后下降的趋势。
注:表中数据为平均值±标准差。
3 结论与讨论
3.设置的问题要有灵活性。同一教学方法可以解决不同的教学内容,不同的教学方法也可以解决相同的教学内容;同一教学方法面对不同的教学对象会产生不同的教学效果,不同的教学方法面对相同的教学对象也会产生不同的教学效果。因此,教学策略的运用要随着问题、目标、内容和教学对象的不同而改变。
数据分析采用SPSS 19.0程序、Microsoft Excel 2013程序。
本研究对湖北海棠在淹水期间的苗高、地径进行了动态观测并对其外部形态进行数字图像记录,发现经淹水后,湖北海棠的高生长在前期受抑制作用不显著,随着淹水时间延长,后期抑制作用较为显著并且此时叶片大多凋落,原因可能是湖北海棠经淹水胁迫后光合能力受限,因此光合产物总量有所下降[23-24],导致湖北海棠生长所需的营养供给不足。而地径在淹水15 d时,处理B的相对生长率开始上升,在淹水35 d时处理B的相对生长率继续上升,且上升至最大(2.7%),原因可能是与湖北海棠为适应淹水胁迫而在茎基部形成通气组织有关[24]。
综上所述,湖北海棠的苗高在淹水胁迫下后期较前期相比受抑制虽较为显著,但总体上不受淹水影响;而地径虽在个别天受抑制,但其余天数受抑制作用不显著,基本上不受淹水影响,因此可以看出湖北海棠具有一定的耐水湿的能力。
植物在淹水胁迫下会导致气孔关闭,光合作用相关酶活性降低,叶绿素发生降解,继而导致叶绿素含量下降,最终植物光合能力的下降[25-26]。
植物的光合色素在光合作用中不仅承担光能的吸收,而且还参与能量的传递与转化,光合色素质量分数的降低将直接影响光合作用的效率[27],在淹水胁迫下,叶绿素的合成与分解被打破,使得叶绿素质量分数降低,此为植物叶片对淹水胁迫的生化响应[28]。研究发现,湖北海棠经淹水后叶绿素相对含量明显降低,而花青素、类胡萝卜素显著升高,由此可见淹水使得叶绿素合成受阻,从而影响叶片的光合作用。在光合作用方面,淹水处理并未显著影响细胞间隙CO2摩尔分数,但显著降低了净光合速率、气孔导度、气孔限制值。除了叶绿素含量降低这一原因,淹水使得根系长时间处于缺氧状态,不能正常的进行新陈代谢以及养分的吸收从而导致湖北海棠光合速率下降,最终光合作用受到抑制,其中气孔限制因素和非气孔因素是其主要原因,根据Farquhar等[25]的观点细胞间隙CO2摩尔分数下降和气孔限制值增大是气孔导度引起光合速率下降的充分条件,本研究中发现淹水胁迫后气孔限制值显著下降,而细胞间隙CO2摩尔分数则与对照差异不显著,由此推测气孔限制因素并不是导致淹水时湖北海棠净光合速率降低的原因,而非气孔限制因素才是导致其净光合速率下降的主要原因之一,这与曹福亮等[19]对乌桕的淹水研究相符。
本研究还发现,淹水后湖北海棠的净光合速率、气孔导度值以及并不是随时间的推移而持续下降,而是大约在在淹水的第10~20 d左右有一定的回升,随后下降。原因可能是湖北海棠在淹水期间产生不定根,使得根内部组织的孔隙度增大,提高了根部对氧气的吸取,改善了有氧环境,使湖北海棠在淹水环境中得以适应[27]。
光合作用是植物生长与代谢等生命活动的基础,植物90%以上的干物质来自光合作用积累的有机物[29]。这说明光合作用和植物的生长密切相关。综上所述,湖北海棠在淹水胁迫后虽然生长、光合能力有所下降,但随着淹水时间的延长,湖北海棠做出适应性响应,各项指标(如净光合速率、气孔导度、光合色素等)变化趋于平稳,因此可看出湖北海棠是一种耐淹能力较强的苗木。
[1] 何秀娟,邱文明,徐育海.湖北海棠观赏型新品系筛选研究[J].湖北林业科技,2014,43(4):5-8.
[2] 罗昌国,渠慎春,张计育,等.湖北海棠MhWRKY40b在几种胁迫下的表达分析[J].园艺学报,2013,40(1):1-9.
[3] 魏要武,向俊,匡仁亮,等.湖北海棠植物资源研究进展[J].园艺与种苗,2012(8):37-40.
[4] 张坤玺.苹果砧木幼苗耐碱性综合评价及营养元素积累的差异分析[D].杨凌:西北农林科技大学,2016.
[5] 张计育.湖北海棠抗病相关基因的克隆及其功能分析[D].南京:南京农业大学,2011.
[6] 陈卫平,冯金侠,孔令娜.八棱海棠和湖北海棠的耐盐性分析[J].湖北农业科学,2011,50(9):1826-1828.
[7] 颜超,王中生,安树青,等.濒危植物银缕梅(Parrotiasubaequalis)不同径级个体的光合能力差异与更新限制[J].生态学报,2008,28(9):4153-4161.
[8] 谭淑端,朱明勇,张克荣,等.植物对水淹胁迫的响应与适应[J].生态学杂志,2009,28(9):1871-1877.
[9] ARMSTRONG W, BRANDLE R, JACKSON M B. Mechanisms of flood tolerance in plants[J]. Plant Biology,2013,43(4):307-358.
[10] 陈鹭真,林鹏,王文卿.红树植物淹水胁迫响应研究进展[J].生态学报,2006,26(2):586-593.
[11] 胡田田,康绍忠.植物淹水胁迫响应的研究进展[J].福建农林大学学报(自然科学版),2005,34(1):18-24.
[12] LORETI E V H, PERATA P. Plant responses to flooding stress[J]. Current Opinion in Plant Biology,2016,33:64-71.
[13] TOUGOU M, HASHIGUCHI A, YUKAWA K, et al. Responses to flooding stress in soybean seedlings with the alcohol dehydrogenase transgene[J]. Plant Biotechnology,2012,29(3):301-305.
[14] 庞宏东,付达夫,胡兴宜,等.淹水胁迫对夹竹桃主要生理生化特性的影响[J].湖北林业科技,2015(4):1-4.
[15] 郁万文,蔡金峰,高长忠.不同桃砧类型对淹水胁迫的生理响应及耐涝性评价[J].中国果树,2016(3):1-6.
[16] ZHAO H, ZHAO Y, ZHANG C, et al. Effect of flooding stress on growth and photosynthesis characteristics ofSalixintegra[J]. Acta Ecologica Sinica,2013,33(3):898-906.
[17] HTAY O, ARAKI T, KUBOTA F. Effects of drought and flooding stresses on growth and photosynthetic activity ofMungbean,Vignaradiata(L.)Wilczekcultivars[J]. Journal of the Faculty of Agriculture Kyushu University,2005,50(2):533-542.
[18] IKEZAWA K, ONJO M, YOSHIDA R, et al. Effects of flooding on photosynthesis in eddo[J]. Japanese Journal of Crop Science,2015,84(2):150-154.
[19] 曹福亮,蔡金峰,汪贵斌,等.淹水胁迫对乌桕生长及光合作用的影响[J].林业科学,2010,46(10):57-61.
[20] 渠慎春,罗昌国,章镇,等.‘湖北海棠’MhWRKY40a基因及其应用[J].南京农业大学学报,2013,36(3):29-35.
[21] MERZLYAK M N, SOLOVCHENKO A E, GITELSON A A. Reflectance spectral features and non-destructive estimation of chlorophyll, carotenoid and anthocyanin content in apple fruit[J]. Postharvest Biology & Technology,2003,27(2):197-211.
[22] SIMS D A, GAMON J A. Relationships between leaf pigment content and spectral reflectance across a wide range of species, leaf structures and developmental stages[J]. Remote Sensing of Environment,2002,81(2/3):337-354.
[23] 杨静,何开跃,李晓储,等.淹水胁迫对两种栎树生长的影响[J].林业科技开发,2008,22(4):34-37.
[23] STRIKER G G, IZAGUIRRE R F, MANZUR M E, et al. Different strategies ofLotusjaponicasLcorniculatusand L.tenuisto deal with complete submergence at seedling stage[J]. Plant Biology,2012,14(1):50.
[24] 潘澜,薛立.植物淹水胁迫的生理学机制研究进展[J].生态学杂志,2012,31(10):2662-2672.
[25] FARQUHAR G D, SHARKEY T D. Stomatal conductance and photosynthesis. annu rev plant physiol plant mol biol[J]. Annual Review of Plant Physiology,1982,33(1):74-79.
[26] ASHRAF M, ARFAN M. GAS exchange characteristics and water relations in two cultivars of Hibiscus esculentus under waterlogging[J]. Biologia Plantarum,2005,49(3):459-462.
[27] 朱向涛,金松恒,哀建国,等.淹水胁迫下江南牡丹生长及光合特性研究[J].广西植物,2016,36(8):956-962.
[28] 张柔,薛立,郭淑红,等.淹水胁迫对南五味子和海南红豆幼苗光合特性的影响[J].湖南林业科技,2012,39(1):17-20.
[29] 刘春风,张往祥,孙垒,等.高温对观赏海棠生长和光合作用的影响[J].南京林业大学学报(自然科学版),2013,37(4):17-22.
EffectsofWaterloggingStressontheGrowthandPhotosynthesisofMalushupehensis(Pamp)Rehd
Zhang Hu
(Jiangsu Vocational and Technical College of Agriculture and Forestry, Jurong 212400, P. R. China); Jiang Wenlong, Wu Qifei, Zhang Wangxiang, Cao Fuliang(Nanjing Forestry University)Journal of Northeast Forestry University,2017,45(12):22-26.
Malushupehensis; Flooding stress; Photosynthesis
1)江苏省科技重点研发计划(BE2016332);江苏省高等职业院校国内高级访问学者计划项目(2015FX026)。
张虎,男,1974年4月生,江苏农林职业技术学院,副教授。E-mail:315725756@qq.com。
2017年3月20日。
潘 华。
S718;Q945
With 1-aMalushupehensisseedling, we studied the effects of flooding stress on the growth and photosynthesis ofM.hupehensis. In the early, the stage of flooding (0-15 d), the inhibition of the growth ofM.hupehensiswas not significant, and the degree of inhibition was significantly increased at the later stage of flooding (15-35 d), and the final seedling height was 10.7%. The inhibitory effect of flooding stress on the diameter ofM.hupehensiswas not significant. Photosynthetic rate, stomatal conductance and stomatal limitation were significantly lower than those of the control during flooding, while intercellular CO2concentration was not significantly affected by flooding. Under the flooding stress, the relative contents of anthocyanins and carotenoids in the leaves ofM.hupehensiswere significantly higher than those of the control, while the relative contents of chlorophyll were significantly lower than those of the control. Therefore,M.hupehensisis a strong water resistance seedlings.
猜你喜欢
农业工程学报(2022年3期)2022-04-16
快乐作文(1.2年级)(2021年2期)2021-09-10
生态学报(2021年6期)2021-04-25
农业工程学报(2021年3期)2021-04-15
节水灌溉(2020年9期)2020-09-16
快乐作文(1.2年级)(2019年12期)2019-09-10
快乐作文(1.2年级)(2019年8期)2019-09-10
新课程·下旬(2017年7期)2017-08-14
传奇故事(破茧成蝶)(2015年5期)2015-02-28
西北林学院学报(2015年5期)2015-01-03
