
表皮生理功能改变在特应性皮炎皮肤瘙痒发生中的作用
2018-01-09 20:53蔄茂强马琳PeterElias胡立志
中华皮肤科杂志 2017年12期
蔄茂强 马琳 Peter M Elias 胡立志
94121旧金山,美国旧金山退伍军人医疗中心、美国北加州研究教育所和加利福尼亚大学旧金山分校医学院皮肤科(蔄茂强、Peter M Elias);首都医科大学附属北京儿童医院皮肤科(马琳);天津医科大学基础学院(胡立志)
·综述·
表皮生理功能改变在特应性皮炎皮肤瘙痒发生中的作用
蔄茂强 马琳 Peter M Elias 胡立志
94121旧金山,美国旧金山退伍军人医疗中心、美国北加州研究教育所和加利福尼亚大学旧金山分校医学院皮肤科(蔄茂强、Peter M Elias);首都医科大学附属北京儿童医院皮肤科(马琳);天津医科大学基础学院(胡立志)
瘙痒是指皮肤对于刺激所导致搔抓反应的欲望,很多系统和皮肤疾病均可导致皮肤瘙痒,根据起源的不同可将瘙痒分为不同的类型。皮肤型瘙痒是由于皮肤病变而导致的瘙痒,是临床最常见的瘙痒类型[1]。皮肤型瘙痒患者往往伴有表皮生理功能(包括表皮通透屏障功能、角质层含水量和表皮酸碱度)异常,如银屑病、湿疹和特应性皮炎。改善表皮功能可能是治疗特应性皮炎患者皮肤瘙痒及相关疾病(包括患者生活和睡眠质量)的有效方法。
一、瘙痒的发生机制和传导通路
引起皮肤瘙痒的主要局部介质包括组胺、P物质、蛋白激酶、白三烯B4、缓激肽、乙酰胆碱、前列腺素E2、5羟色胺和白细胞介素31(IL⁃31)等。这些皮肤局部介质主要来源于角质形成细胞和炎症细胞。如T细胞可产生IL⁃31,IL⁃31在银屑病、特应性皮炎和结节性痒疹等瘙痒性皮肤病中的表达明显升高;肥大细胞等可产生白三烯B4,特应性皮炎患者皮肤中白三烯B4的表达明显升高;嗜碱性粒细胞和肥大细胞可合成组胺和蛋白激酶。角质形成细胞可以产生组胺、前列腺素E2和缓激肽。此外,C神经纤维末稍受刺激后可释放P物质。引起皮肤局部瘙痒的主要介质及介质的主要来源见表1[2⁃4]。介质不仅可由一种细胞分泌,也可由多种细胞相互作用产生,一些炎症介质既可以直接引起瘙痒,也可以刺激其他炎症细胞产生相应的致痒介质。如角质形成细胞可分泌脂质运载蛋白2、组胺以及炎症细胞因子[5⁃6]。此外,虽然角质形成细胞本身不能合成白三烯B4,但是中性粒细胞可以将白三烯A4转运到角质形成细胞内,在白三烯A4水解酶的作用下角质形成细胞将白三烯A4转化成白三烯B4,产生致痒作用。综上所述,皮肤瘙痒(尤其是炎症性皮肤病所致的瘙痒)可由多种细胞多介质共同作用而引起。

表1 引起皮肤局部瘙痒的主要介质及其来源
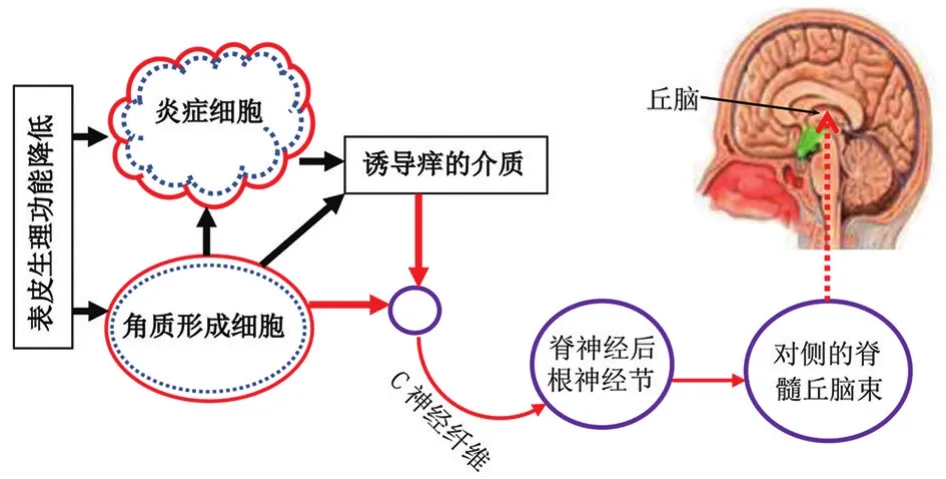
图1 痒的产生及其主要传导通路 角质形成细胞和炎症细胞均可分泌炎症细胞因子激活表皮C神经纤维的受体,经由脊神经节-脊髓丘脑束将信号传至丘脑而引起痒感。角质形成细胞既可以刺激炎症细胞分泌炎症细胞因子致痒,也可以直接作用于表皮C神经纤维而致痒
皮肤痒感的传导神经纤维主要是慢传导无髓鞘C纤维(包括机械刺激不敏感型和机械及热刺激敏感型)和有髓鞘Aδ纤维[7⁃9]。由于组胺和非组胺类介质分别激活不同的神经元,所以,当这些介质在皮肤局部增多时,可通过组胺和非组胺途径激活位于真表皮交界处的神经末梢,经脊丘束传导至丘脑而导致痒感(图1)。当然这种分类并非绝对,大约有2%的神经元对组胺和非组胺刺激均敏感[10]。组胺途径可分为瞬时受体电位香草酸亚型1(transient receptor potential vanilloid receptor 1,TRPV1)阳性和TRPV1阴性途径。组胺受体(Hr)系G蛋白偶联受体,其中,H1r、H3r和H4r均表达于支配感觉神经元。当组胺与相应感觉神经元的受体结合后,激活磷脂酶A2和脂加氧酶,继而导致钙离子内流和激活TRPV1,产生内流电流,最终导致痒感。在培养的神经细胞中,如果仅有TRPV1或组胺受体的表达,用TRPV1阻滞剂能减轻组胺所致的内向电流,这一途径称为组胺-TRPV1阳性途径。在组胺-TRPV1阳性途径中,组胺与H4r结合后,可激活磷脂酶C生成三磷酸肌醇(IP3)和甘油二酯[11]。IP3引起钙离子内流并激活TRPV1,而甘油二酯能直接激活TRPV1,导致痒感。还有一类亚型C神经纤维同时表达H1r和磷脂酶Cβ3,组胺可以通过磷脂酶Cβ3⁃TRPV1信号通路引起瘙痒[12]。抗组胺制剂能抑制激活蛋白激酶激活受体1(protease activated receptor 1,PAR1)和 PAR3所致的瘙痒,提示组胺-PAR通路在皮肤瘙痒中的作用[13]。通常经组胺途径引起瘙痒的神经纤维属于机械刺激不敏感型C纤维,也有部分神经纤维对机械刺激不敏感而对组胺敏感,组胺激活这类神经纤维产生的痒感属于TRPV1阴性途径。非组胺途径主要是通过PAR介导激活对机械刺激敏感C神经纤维而导致痒感。PAR也属于G蛋白偶联受体,共有4个亚型(PAR1~4),都表达于神经元[14⁃15]。在痒感的形成过程中,不同的致痒介质激活不同的PAR亚型,如,Mucunain(刺毛黎豆的成分)通过激活PAR2与PAR4致痒;丝氨酸蛋白酶(serine protease,主要来自角质形成细胞)和类胰蛋白酶(tryptase,主要来自肥大细胞)则是通过PAR2致痒。在特应性皮炎中,主要是丝氨酸蛋白酶-PAR2通路参与痒感的发生。此外,有些感觉神经元能同时表达IL⁃31受体、TRPV1和瞬时受体电位锚蛋白亚型1(transient receptor potential cation channel ankyrin subtype 1,TRPA1),IL⁃31是通过TRPV1和TRPA1引起瘙痒[3]。而具体到特应性皮炎,其皮肤瘙痒的发生是多介质介导、诸多信号传递通路参与的复杂过程。

图2 表皮功能改变在特应性皮炎瘙痒发生中的作用机制 主要表皮功能改变包括:表皮通透屏障功能降低、角质层含水量减少和角质层pH升高。表皮通透屏障功能降低一方面通过促进角质形成细胞炎症细胞因子释放直接导致瘙痒,另一方面炎症细胞因子促进组织肥大细胞浸润、炎症因子分泌和蛋白激酶活化而导致瘙痒;角质层含水量减少能导致炎症细胞浸润和和炎症细胞因子释放而导致瘙痒;角质层pH升高可通过激活神经纤维的PAR2受体而导致瘙痒。PAR2:蛋白激酶激活受体2
二、特应性皮炎患者表皮生理功能改变对瘙痒发生的影响
特应性皮炎患者的表皮生理功能改变主要包括表皮通透屏障功能降低、角质层含水量减少和角质层pH增高。这些功能的改变不仅能进一步加重疾病,还能通过多种信号传导通路引起皮肤瘙痒(图2)。
1.表皮通透屏障功能:表皮通透屏障功能减低既是特应性皮炎的发病诱因,也是结果。表皮通透屏障功能降低使致敏物更易进入皮肤导致皮炎,当皮炎发生后,表皮通透屏障功能进一步降低。特应性皮炎的严重程度与透皮失水率呈正相关,而透皮失水率与瘙痒程度呈正相关。动物实验证明,动物的搔抓次数与透皮失水率成正比。由此提示,表皮通透屏障功能降低与特应性皮炎瘙痒的发生有关。首先,表皮通透屏障功能减低能刺激表皮合成和释放炎症因子,诱导朗格汉斯细胞、肥大细胞等炎症细胞浸润[16]。其次,表皮通透屏障功能减低能促进T细胞浸润、增殖和成熟。这样角质形成细胞和炎症细胞产生的各种致痒介质可通过多种途径刺激神经纤维致痒。此外,表皮通透屏障功能减低可增加表皮丝氨酸蛋白激酶的表达和活性,继而激活PAR2,最终致痒。PAR2能促进角质形成细胞产生胸腺间质淋巴细胞生成素(thymic stromal lymphopoietin,TSLP),后者经TRPA1通路致痒[17⁃18]。表皮通透屏障功能降低能使表皮神经生长因子mRNA的表达增加20~30倍,神经生长因子促进神经增生,最终导致表皮神经纤维增多,也会增加皮肤对致痒介质的敏感性。
2.角质层酸碱度:角质层酸碱度对表皮功能具有重要调节作用。角质层pH值升高可延缓表皮通透屏障功能的恢复和诱发、加重特应性皮炎[19];降低角质层pH不仅有利于表皮通透屏障功能的恢复,还可以预防和治疗特应性皮炎[20]。而特应性皮炎可导致角质层pH升高。动物试验证明,pH升高可增加表皮激活胰肽释放酶相关酶5的表达[21],后者经PAR2⁃TSLP通路不仅使特应性皮炎加重,而且可通过TRPA1引起瘙痒[17⁃18],同时,激活PAR2本身也能致痒。角质层pH升高有利于金黄色葡萄球菌生长[22],由金黄色葡萄球菌产生的δ毒素能刺激肥大细胞,后者释放组胺和细胞炎症因子,导致疾病加重和瘙痒。特应性皮炎患者用酸性水每日洗澡2次,平均60 d后,70%以上的患者皮肤金黄色葡萄球菌减少,痒感减轻[23]。这些证据表明,特应性皮炎角质层pH升高与瘙痒的发生有关。
3.角质层含水量:皮肤干燥与多种原因引起的皮肤瘙痒有关。如老年瘙痒、血液透析和冬季瘙痒症都与皮肤干燥有关[22]。而角质层含水量降低是特应性皮炎患者表皮生理功能改变的特点之一,其导致或加重瘙痒主要通过下列途径:①增加表皮炎症细胞因子如肿瘤坏死因子α、环氧合酶2、IL⁃1β及IL⁃8等的表达[24],这些炎症因子除通过刺激细胞分泌致痒介质外,还能加重5羟色胺导致的痒;②促进皮肤炎症细胞浸润和肥大细胞释放组胺;③增加表皮神经生长因子的表达和表皮神经纤维的密度,继而增加皮肤对致痒介质的敏感性[25];局部外涂保湿剂能防止由于皮肤干燥所导致的表皮神经生长因子表达和神经纤维密度增加,这可能是保湿剂止痒的机制之一;④增加TRPV1阳性以及TRPV1与TRPA1阳性感觉神经元PAR2和缓激肽受体的表达[26]。由于TRPA1敲除的小鼠对皮肤干燥所导致的瘙痒明显减轻,提示TRPA1在介导皮肤干燥所导致瘙痒中发挥重要作用[26]。在临床上,改善角质层含水量能缓解与皮肤干燥所导致的瘙痒,如晚期肾病、老年瘙痒和特应性皮炎[27⁃28]。
综上所述,皮肤瘙痒是由于致痒介质作用于皮肤感觉神经末梢而导致的搔抓欲望。特应性皮炎表皮生理功能改变导致皮肤感觉神经纤维密度增加,致痒介质及其受体表达增多可加重瘙痒。因此,在防治伴有表皮生理功能异常的皮肤瘙痒时,改善表皮生理功能是值得一试的方法之一。
[1]Garibyan L,Rheingold CG,Lerner EA.Understanding the pathophysiology of itch[J].Dermatol Ther,2013,26(2):84⁃91.DOI:10.1111/dth.12025.
[2]Shi X,Wang L,Clark JD,et al.Keratinocytes express cytokines and nerve growth factor in response to neuropeptide activation of the ERK1/2 and JNK MAPK transcription pathways[J].Regul Pept,2013,186:92⁃103.DOI:10.1016/j.regpep.2013.08.001.
[3]Cevikbas F,Wang X,Akiyama T,et al.A sensory neuron⁃expressed IL⁃31 receptor mediates T helper cell⁃dependent itch:involvement of TRPV1 and TRPA1[J].J Allergy Clin Immunol,2014,133(2):448⁃460.DOI:10.1016/j.jaci.2013.10.048.
[4]Lee CH,Hong CH,Yu WT,et al.Mechanistic correlations between two itch biomarkers,cytokine interleukin⁃31 and neuro⁃peptide β⁃endorphin,via STAT3/calcium axis in atopic dermatitis[J].Br J Dermatol,2012,167(4):794⁃803.DOI:10.1111/j.1365⁃2133.2012.11047.x.
[5]Shao S,Cao T,Jin L,et al.Increased lipocalin⁃2 contributes to the pathogenesis of psoriasis by modulating neutrophil chemo⁃taxis and cytokine secretion[J].J Invest Dermatol,2016,136(7):1418⁃1428.DOI:10.1016/j.jid.2016.03.002.
[6]Shimizu K,Andoh T,Yoshihisa Y,et al.Histamine released from epidermal keratinocytes plays a role in α⁃melanocyte⁃stimulating hormone⁃induced itching in mice[J].Am J Pathol,2015,185(11):3003⁃3010.DOI:10.1016/j.ajpath.2015.07.015.
[7]Johanek LM,Meyer RA,Hartke T,et al.Psychophysical and physiological evidence for parallel afferent pathways mediating the sensation of itch[J].J Neurosci,2007,27(28):7490⁃7497.DOI:10.1523/JNEUROSCI.1249⁃07.2007.
[8]Schmelz M,Schmidt R,Weidner C,et al.Chemical response pattern of different classes of C⁃nociceptors to pruritogens and algogens[J].J Neurophysiol,2003,89(5):2441 ⁃2448.DOI:10.1152/jn.01139.2002.
[9]Ringkamp M,Schepers RJ,Shimada SG,et al.A role for nociceptive,myelinated nerve fibers in itch sensation[J].J Neurosci,2011,31(42):14841 ⁃14849.DOI:10.1523/JNEUROSCI.3005⁃11.2011.
[10]Davidson S,Zhang X,Khasabov SG,et al.Pruriceptive spinotha⁃lamic tract neurons:physiological properties and projection targets in the primate[J].J Neurophysiol,2012,108(6):1711 ⁃1723.DOI:10.1152/jn.00206.2012.
[11]Han SK,Mancino V,Simon MI.Phospholipase C beta 3 mediates the scratching response activated by the histamine H1 receptor on C⁃fiber nociceptive neurons[J].Neuron,2006,52(4):691⁃703.DOI:10.1016/j.neuron.2006.09.036
[12]Tsujii K,Andoh T,Lee JB,et al.Activation of proteinase⁃activated receptors induces itch⁃associated response through histamine⁃dependent and ⁃independent pathways in mice[J].J Pharmacol Sci,2008,108(3):385 ⁃388.DOI:10.1254/jphs.08200SC.
[13]Jian T,Yang N,Yang Y,et al.TRPV1 and PLC participate in histamine H4 receptor⁃induced itch[J/OL].Neural Plast,2016,2016:1682972[2016⁃11⁃14].https://www.hindawi.com/journals/np/2016/1682972/.DOI:10.1155/2016/1682972.
[14]Funahashi Y,Namba T,Fujisue S,et al.ERK2⁃mediated pho⁃sphorylation of Par3 regulates neuronal polarization [J].J Neurosci,2013,33(33):13270⁃13285.DOI:10.1523/JNEUROSCI.4210⁃12.2013.
[15]Chen D,Wang Z,Zhang Z,et al.Capsaicin up⁃regulates protease⁃activated receptor⁃4 mRNA and protein in primary cultured dorsal root ganglion neurons[J].Cell Mol Neurobiol,2013,33(3):337⁃346.DOI:10.1007/s10571⁃012⁃9899⁃y.
[16]Lin TK,Man MQ,Santiago JL,et al.Topical antihistamines display potent anti⁃inflammatory activity linked in part to enhanced permeability barrier function[J].J Invest Dermatol,2013,133(2):469⁃478.DOI:10.1038/jid.2012.335.
[17]Moniaga CS,Jeong SK,Egawa G,et al.Protease activity enhances production ofthymic stromallymphopoietin and basophil accumulation in flaky tail mice[J].Am J Pathol,2013,182(3):841⁃851.DOI:10.1016/j.ajpath.2012.11.039.
[18]Wilson SR,Thé L,Batia LM,et al.The epithelial cell⁃derived atopic dermatitis cytokine TSLP activates neurons to induce itch[J].Cell,2013,155(2):285⁃295.DOI:10.1016/j.cell.2013.08.057.
[19]Sakai T,Hatano Y,Zhang W,et al.Defective maintenance of pH of stratum corneum is correlated with preferential emergence and exacerbation of atopic⁃dermatitis⁃like dermatitis in flaky⁃tail mice[J].J Dermatol Sci,2014,74(3):222 ⁃228.DOI:10.1016/j.jdermsci.2014.01.012.
[20]Lee HJ,Yoon NY,Lee NR,et al.Topical acidic cream prevents the development of atopic dermatitis⁃and asthma⁃like lesions in murine model[J].Exp Dermatol,2014,23(10):736⁃741.DOI:10.1111/exd.12525.
[21]Jang H,Matsuda A,Jung K,et al.Skin pH is the master switch of kallikrein 5⁃mediated skin barrier destruction in a murine atopic dermatitis model[J].J Invest Dermatol,2016,136(1):127⁃135.DOI:10.1038/JID.2015.363.
[22]Nakamura Y,Oscherwitz J,Cease KB,et al.Staphylococcusδ⁃toxin induces allergic skin disease by activating mast cells[J].Nature,2013,503(7476):397⁃401.DOI:10.1038/nature12655.
[23]Garibyan L,Chiou AS,Elmariah SB.Advanced aging skin and itch:addressing an unmet need[J].Dermatol Ther,2013,26(2):92⁃103.DOI:10.1111/dth.12029.
[24]Peng XY,Huang Y,Wang XL,et al.Adrenergic β2 ⁃receptor mediates itch hypersensitivity following heterotypic chronic stress in rats[J].Neuroreport,2015,26(17):1003⁃1010.DOI:10.1097/WNR.0000000000000458.
[25]Kamo A,Tominaga M,Taneda K,et al.Neurotropin inhibits the increase in intraepidermal nerve density in the acetone⁃treated dry⁃skin mouse model[J].Clin Exp Dermatol,2013,38(6):665⁃668.DOI:10.1111/ced.12100.
[26]Wilson SR,Nelson AM,Batia L,et al.The ion channel TRPA1 is required for chronic itch[J].J Neurosci,2013,33(22):9283 ⁃9294.DOI:10.1523/JNEUROSCI.5318⁃12.2013.
[27]Cristaudo A,Francesconi L,Ambrifi M,et al.Efficacy of an emollient dermoprotective cream in the treatment of elderly skin affected by xerosis[J].G Ital Dermatol Venereol,2015,150(3):297⁃302.
[28]Hon KL,Pong NH,Wang SS,et al.Acceptability and efficacy of an emollient containing ceramide⁃precursor lipids and moisturi⁃zing factors for atopic dermatitis in pediatric patients[J].Drugs R D,2013,13(1):37⁃42.DOI:10.1007/s40268⁃013⁃0004⁃x.
蔄茂强,Email:mqman@hotmail.com;胡立志,Email:lizhihuleopard@yahoo.com
国家自然科学基金(81573075、81301360);天津医科大学基金(2013KY06)
10.3760/cma.j.issn.0412⁃4030.2017.12.024
志谢本文部分工作是在美国旧金山退伍军人医疗中心完成
2016⁃12⁃02)
朱思维 颜艳)
猜你喜欢
中国现代医生(2022年19期)2022-08-25
皮肤病与性病(2021年3期)2021-07-30
智慧健康(2021年33期)2021-03-16
昆明医科大学学报(2020年12期)2021-01-26
中华养生保健(2020年10期)2021-01-18
作文周刊·小学二年级版(2021年40期)2021-01-05
中国合理用药探索(2020年9期)2020-10-13
伴侣(2020年4期)2020-04-27
伴侣(2020年2期)2020-04-13
健康必读(2016年10期)2016-11-14
