
辣椒热激转录因子家族在驯化过程中的演变
2018-01-23 08:42龚记熠孙媛媛
西北大学学报(自然科学版) 2018年1期
李 菲,龚记熠,孙媛媛,乙 引
(贵州师范大学 生命科学学院/贵州省植物生理与发育调控重点实验室, 贵州 贵阳 550001)
不能移动的植物生存在变动的环境中,需要应对各种不利的环境条件。目前针对植物抗性的研究,往往聚焦于某一单一逆境。转录组学分析发现,61%的双重胁迫诱导的基因在任何一个单一胁迫环境下都不会诱导表达[1]。因此,有必要研究植物响应多重胁迫的机制[2-3]。
植物应对多种逆境的方法之一是利用多效热激转录因子(Heat Shock Transcription Factors,HSFs)——热激蛋白/分子伴侣(heatshock protein/chaperone)信号传导和基因调控网络。逆境影响植物正常生长的分子机制,是阻碍细胞内蛋白质正常进行生理功能。蛋白质变性是任何逆境都会引发的后果。所以,任何会引起蛋白质错误折叠的逆境,都需要植物基因组中热激转录因子(HSFs)信号传导和基因调控网络来应对[4]。热激转录因子(HSFs)信号传导途径的作用不仅限于热应激响应,而且还涉及多种胁迫,如冷、渗透、干旱、盐、紫外线、高光、氧化和病原体侵染胁迫[5]。
HSF存在于所有真核生物中,但与酿酒酵母中的1个HSF或人类中7个HSFs基因相比,植物基因组中普遍存在大量HSFs基因(大豆中38个,水稻中25个,拟南芥21个)[6]。植物中HSF家族的多样性造成了研究的困难。然而,表达模式、基因突变和过表达研究显示了拟南芥HSFs家族基因在抗多种逆境胁迫和发育中的独特和重叠的功能[7]。
热激转录因子(HSFs)分为A,B和C3类[8-10]。大量的HSFs成员基因及其以异源多聚体的形式发挥功能的特征,使得特定基因的功能研究非常困难。多个HSFs基因在植物干旱、盐、强光、冷胁迫等的信号传导和基因表达调控中起作用。它们可以被ABA依赖性或非依赖的信号通路诱导。HSFA3是DREB2A转录因子)的靶点,由多种胁迫如干旱、盐、热和冷以及氧化胁迫、紫外光UVB和伤害诱导[11-12]。AtHSFA6a的启动子包含两个ARE(ABA反应元件),并在体外结合3种ABA响应转录因子:AREB1,ABF3和AREB3[13]。过表达AtHSFA6a的植物表现出抗盐胁迫和抗旱能力。B类和C类HSFs基因缺少A类HSFs基因调节转录活性所必需的结构域AHA (aromatic hydrophobic acidic),因此被认为是抑制性转录因子。
在非模式植物中,HSFs转录因子的功能也有研究[14]。辣椒营养丰富,是我国西南省份重要的经济作物。利用重要的栽培辣椒品种遵辣一号(C.apsicumannuumLZunla1),及其野生祖先品系Chiltepin(C.annuumvar.glabriusculum)的基因组测序和注释结果。本研究分析了野生辣椒和栽培辣椒基因组中HSFs基因家族的成员组成、系统分类和可能功能,以及驯化过程中辣椒基因组中HSFs基因家族发生的演变。本研究为辣椒品系应对多种逆境的机制研究和品种改良提供了基因资源。
1 材料与方法
1.1 拟南芥HSFs基因家族序列和辣椒基因组序列来源
拟南芥HSFs基因家族序列来源于拟南芥数据库TAIR。栽培辣椒遵辣一号和其野生祖先品系Chiltepin的基因组序列及其注释信息可以从NCBI Genome数据库下载。
1.2 辣椒基因组中HSFs基因的检索和筛选
辣椒基因组中HSFs基因的检索工具是本地BLAST软件。将下载到的栽培辣椒遵辣一号和其野生祖先品系Chiltepin的基因组序列及其注释信息,构建本地核苷酸和氨基酸数据库。以拟南芥HSFs家族全部序列信息为检索序列,进行本地BLAST。保留检索到的E值小于0001的序列。
下一步对这些序列进行结构域分析。结构域分析是在Pfam在线蛋白质结构域数据库中,进行这些序列的结构域检索。保留含有HSFs特征性结构域:HSF-DNAbind结构域的序列,用于后续分析。
1.3 栽培辣椒和野生辣椒HSFs基因家族的染色体定位比较
栽培辣椒和野生辣椒HSFs基因家族成员的染色体定位信息可以根据基因序列号从基因组序列信息中查询到。根据基因的染色体定位信息,使用MapInspect软件分别构建出栽培辣椒和野生辣椒HSFs基因家族成员的染色体定位图谱,并进行比对和解读。
1.4 辣椒基因组中HSFs基因的系统进化分析
对拟南芥、栽培辣椒遵辣一号和野生辣椒品系Chiltepin 全部HSFs基因序列进行序列比对和系统进化树构建,使用的软件是MEGA70。序列比对使用的算法是Clustal W。系统发育树构建使用的算法是Neighbor Joining。
2 结果与讨论
2.1 辣椒基因组中HSFs基因的鉴定和特征初步分析
在西南各省广泛种植的栽培辣椒品种遵辣一号(C.apsicumannuumLZunla1),和其野生祖先品系Chiltepin(C.annuumvar.glabriusculum)的基因组测序已经完成,序列及注释信息均公开(http://peppersequencegenomicscn/page/species/indexjsp)。使用拟南芥HSFs基因家族的基因序列,可以从辣椒基因组中挖掘出其中的HSFs基因。
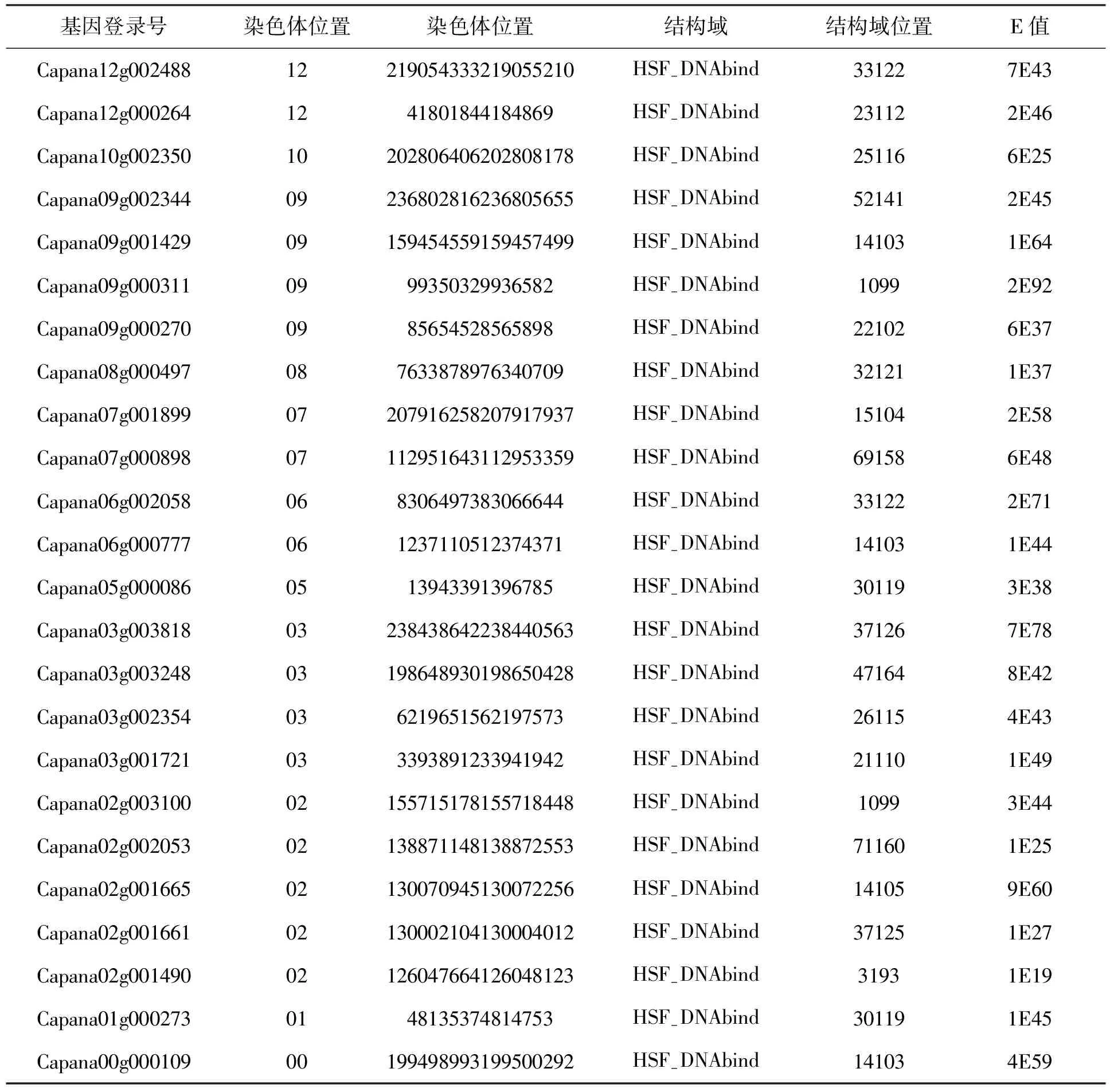
表1 遵辣一号(C. apsicum annuum LZunla1)基因组中HSFs基因信息列表Tab.1 The HSFs genes in the C. apsicum annuum L Zunla1 genome
在栽培辣椒遵辣一号基因组中共鉴定出25个HSFs基因,其野生祖先品系Chiltepin中鉴定出26个HSFs基因。这一HSFs基因家族成员数目与其他植物(拟南芥21,水稻25个,大豆38个)相差不多(Fujimoto and Nakai 2010),但比其他生物,如动物和酵母多。这应该与植物必须静止应对多种逆境胁迫有关。辣椒基因组经过重复序列的扩增,比其他植物大很多(辣椒3364Mbp;拟南芥125Mbp;水稻466Mbp; 大豆1100Mbp)。但是,基因组中HSFs功能基因的数量,辣椒并没有比其他植物更多。
2.2 栽培辣椒和野生辣椒品系基因组中HSFs基因的染色体定位
栽培辣椒品种遵辣一号(C.apsicumannuumLZunla1),和其野生祖先品系Chiltepin(C.annuumvar.glabriusculum)的农艺性状,包括株高、株型、果实大小等有巨大的差异,在抗性方面也有不同的表现。从HSFs基因染色体位置看,与其野生祖先品系Chiltepin比较,遵辣一号HSFs基因家族基本一致。大部分HSFs基因在驯化过程中没有发生染色体位置变化。在遵辣一号品种一号和七号染色体上,分别丢失了1个HSFs基因(Capang01g003145和Capang07g001960)。遵辣一号基因组中有1个HSF基因,在注释过程中未能定位到任何一个染色体上(Capana00g000109)。这个基因和Chiltepin野生辣椒品系的Capang04g002355基因在系统发育树中聚集在一个分支,与Chiltepin基因组中Capang01g003145和Capang07g001960并没有相近的亲缘关系。
2.3 栽培辣椒和野生辣椒品系基因组中HSFs基因的系统发育分析
对于HSFs基因家族的功能,研究的最透彻的还是拟南芥。将拟南芥、栽培辣椒遵辣一号和野生辣椒Chiltepin的全部HSFs基因的氨基酸序列进行系统进化分析,如图2。
与拟南芥一样,辣椒HSFs基因家族分成了A/B/C3个亚家族。从系统发育树中,可以清晰地看到辣椒HSFs基因在拟南芥中的同源基因。遵辣一号的HSFs基因,一般与Chiltepin的HSFs基因一一对应,位于同一分支。少数几个例外包括:Capana00g000109和Chiltepin野生辣椒品系的Capang04g002355在系统发育树中聚集在一个分支;Chiltepin野生辣椒品系的Capang07g001960基因在遵辣一号中没有对应基因;遵辣一号的Capana08g000497 与Chiltepin野生辣椒品系的Capang01g003145基因聚集在一个分支。

图1 栽培辣椒遵辣一号” Zunla1”和野生辣椒品系” Chiltepin”基因组中HSFs基因的染色体定位图谱Fig.1 Chromosomal location map of HSFs gene in the genome of the cultivated pepper "Zunla1" and its wild ancestor "Chiltepin"
2.4 栽培辣椒和野生辣椒品系基因组中HSFs基因的可能功能
HSFs基因家族成员,尤其是HSFAs亚家族的成员,功能研究的报道已经有很多。系统发育分析揭示了与拟南芥HSFAs基因同源的辣椒基因。这为对辣椒HSFs基因进行功能研究提供了很好的线索。
在拟南芥中,热激转录因子HSFA1abd和e组成型表达,负责触发热应激反应(HSR,heat stress response)[11]。这些转录因子与热激蛋白启动子区域的热应激因子(HSE heat stress elements)元件结合以激活热激蛋白HSP的转录。此外,这些基因也可激活其他热激转录因子,包括HSFA2、HSFA3和HSFA7a的表达,赋予植株长时间的胁迫抗性[15]。这些信号通路中,HSFA2处于重要的中心位置。已经充分证明,HSFA2的过表达可增强植株对热、缺氧、盐、渗透胁迫、强光和氧化胁迫的抵抗力[16]。
与拟南芥HSFA2基因同源的是遵辣一号的Capana08g000497 与Chiltepin野生辣椒品系的Capang01g003145基因。遵辣一号的Capana08g000497 与Chiltepin的Capang01g003145两个基因相似度为100%。与拟南芥HSFA2基因相似度为89%。鉴于HSFA2基因对于拟南芥多种抗性的重要作用,研究遵辣一号的Capana08g000497 与Chiltepin的Capang01g003145基因在辣椒中的功能,具有重要的意义。
这一重要的抗逆基因,在驯化过程中,由Chiltepin一号染色体移动到遵辣一号的八号染色体。这一基因移位发生的时间和机制、基因移位对该基因表达的影响,以及对栽培辣椒抗逆性的影响,是研究栽培辣椒抗逆性的关键问题。

图2 栽培辣椒遵辣一号” Zunla1”和野生辣椒品系” Chiltepin”基因组中HSFs基因与拟南芥HSFs基因家族的系统进化分析Fig.2 Phylogenetic analysis of HSFs gene in the genome of Arabidopsis, the cultivated pepper "Zunla1" and its wild ancestor "Chiltepin"
3 结 论
气候灾害环境给农作物生产带来严重的风险,极大的危害农业经济效益的稳定性。目前对植物抗逆的研究,主要集中在对单个逆境的研究。这很难解决实际生产中遇到的多种逆境同时发生的情况。植物基因组中的热激转录因子——热激蛋白信号传导和基因表达调控途径,是植物在各种胁迫环境下,均会激活以便维持细胞内蛋白质正常生理功能的重要抗逆机制。本文以研究相对透彻的拟南芥HSFs基因家族已知信息为基础,挖掘辣椒栽培品种遵辣一号(CapsicumannuumLZunla1),及其野生祖先品系Chiltepin(C.annuumvar.glabriusculum)基因组中HSFs基因,并对这些基因进行系统的比对和功能分析。
尽管辣椒基因组远大于拟南芥,其基因组中HSFs基因的数目并没有更多。对辣椒和其他茄科植物,如番茄、茄子等的基因组进行比较,发现辣椒基因组中重复序列发生了大规模的扩增,但是基因数目并没有增加[17]。对辣椒基因组中HSFs基因家族的分析,为此论断提供一个新的例证。
在栽培辣椒和其野生祖先辣椒品系基因组中,HSFs基因家族比较保守。栽培辣椒遵辣一号(C.apsicumannuumLZunla1)基因组中,HSFs基因家族成员的数目、染色体位置和序列,与其野生祖先大体上保持了一致。但是,可提高拟南芥对多种逆境抗性的重要基因,HSFA2,在遵辣一号及其野生祖先Chiltepin中的同源基因发生了移位。这一基因移位对于栽培辣椒与其野生祖先抗逆性的差异之间的关联,是值得研究的问题。本研究为分析辣椒HSFs基因家族成员的功能提供了有价值的信息,为辣椒品种多种抗逆能力的改善,积累了基因资源。
[1] RASMUSSEN S, BARAHarah P, SUAREZRODRIGUEZ M C, et al. Transcriptome responses to combinations of stresses in arabidopsis[J]. Plant Physiology, 2013, 161(4):1783-1794.
[2] MITTLER R, BLUMWALD E. Genetic engineering for modern agriculture: Challenges and perspectives[J]. Annual Review of Plant Biology, 2010, 61(1):443-462.
[3] NOBUHIRO SUZUKI, RIVERO R M, VLADIMIR S, et al. Abiotic and biotic stress combinations[J]. New Phytologist, 2014, 203(1):32-43.
[4] CAREY C C, GORMAN K F, RUTHERFORD S. Modularity and intrinsic evolvability of Hsp90-buffered change[J]. Plos One, 2006, 1(1):76.
[5] SWINDELL W R, HUEBNER M, WEBER A P. Transcriptional profiling of Arabidopsis heat shock proteins and transcription factors reveals extensive overlap between heat and non-heat stress response pathways[J]. Bmc Genomics, 2007, 8(1):125.
[6] FUJIMOTO M, NAKAI A. The heat shock factor family and adaptation to proteotoxic stress[J].Febs Journal, 2010, 277(20):4112-4125.
[7] VON K P, SCHARF K D, NOVER L. The diversity of plant heat stress transcription factors[J].Trends in Plant Science, 2007, 12(10):452-457.
[8] DORING P. Arabidopsis and the heat stress transcription factor world: How many heat stress transcription factors do we need?[J]. Cell Stress & Chaperones, 2001, 6(3):177-189.
[9] SCHARF K D, BERBERICH T, EBERSBERGER I, et al. The plant heat stress transcription factor (Hsf) family:Structure, function and evolution.[J]. Biochimica et Biophysica Acta, 2012, 1819(2):104-119.
[10] GUO M, LIU J H, MA X, et al. The plant Heat Stress Transcription factors (HSFs): Structure, regulation, and function in response to abiotic stresses[J]. Frontiers in Plant Science, 2016, 7(273):114.
[11] YOSHIDA T, SAKUMA Y, TODAKA D, et al. Functional analysis of an Arabidopsis, heat-shock transcription factor HsfA3, in the transcriptional cascade downstream of the DREB2A stress-regulatory system[J]. Biochemical & Biophysical Research Communications, 2008, 368(3):515-521.
[12] DEBBIE WINTER, BEN VINEGAR, HARDEEP NAHAL, et al. An "Electronic Fluorescent Pictograph" browser for exploring and analyzing Large-Scale biological data sets[J]. PloS one, 2007, 2(8):e718.
[13] HWANG S M, KIM D W, WOO M S, et al. Functional characterization of Arabidopsis HsfA6a as a heat-shock transcription factor under high salinity and dehydration conditions[J]. Plant Cell & Environment, 2014, 37(5):1202-1222.
[14] MISHRA S K, TRIPP J, WINKELHAUS S, et al. In the complex family of heat stress transcription factors, HsfA1 has a unique role as master regulator of thermotolerance in tomato[J]. Genes & Development, 2002, 16(12):1555-1567.
[15] NISHIZAWAYOKOI A, NOSAKA R, HAYASHI H, et al. HsfA1d and HsfA1e involved in the transcriptional regulation of HsfA2 function as key regulators for the Hsf signaling network in response to environmental stress[J]. Plant & Cell Physiology, 2011, 52(5):933-945.
[16] DAISUKE OGAWA, KAZUO YAMAGUCHI, TAKUMI NISHIUCHI. High-level overexpression of the Arabidopsis HsfA2 gene confers not only increased themotolerance but also salt/osmotic stress tolerance and enhanced callus growth[J]. Journal of Experimental Botany, 2007, 58(12):33-73.
[17] QIN C, YU C, SHEN Y, et al. Whole-genome sequencing of cultivated and wild peppers provides insights into Capsicum domestication and specialization.[J]. Proceedings of the National Academy of Sciences of the United States of America, 2014, 111(14):5135-5140.
猜你喜欢
今日农业(2021年11期)2021-08-13
今日农业(2020年14期)2020-12-14
中国生殖健康(2020年4期)2020-12-09
中西医结合肝病杂志(2020年2期)2020-10-27
山东林业科技(2019年2期)2019-06-03
云南农业科技(2019年2期)2019-04-04
中成药(2018年7期)2018-08-04
上海农业学报(2017年3期)2017-04-10
红领巾·探索(2015年9期)2015-09-10
天然产物研究与开发(2014年6期)2014-04-27
