
不同年限设施菜地番茄细胞壁果胶Cd累积的研究
2018-03-01 08:56郭军康任心豪徐慧荟李艳萍
农业环境科学学报 2018年1期
郭军康,周 冉,任心豪,魏 婷,徐慧荟,李艳萍,吕 欣
(陕西科技大学 环境科学与工程学院,西安 710021)
随着我国设施农业的迅猛发展,全国设施菜地生产面积快速扩大。截止2013年底,我国设施蔬菜生产面积近370万hm2,设施蔬菜已成为现代农业的重要组成部分。设施菜地常年处于半封闭状态,种植强度大,化肥、农药等农用品投入量大,高投入高产出的生产模式和管理方式导致一系列土壤环境问题,城市郊区污水引灌进一步加剧土壤重金属污染问题,影响蔬菜质量安全。《全国土壤污染状况调查公报》表明,我国土壤环境状况总体不容乐观,耕地土壤环境质量堪忧,其中镉(Cd)污染超标严重,其点位超标率达7.0%[1]。Cd在土壤中易累积,难降解,且迁移性较强。镉是植物生长的非必需元素,当Cd含量超标时,会抑制植物根系生长、阻碍养分运输和吸收,同时还会影响光合色素合成,酶活性改变,使得植物光合作用减缓,生物量降低,作物减产[2-4]。
细胞壁是重金属进入植物体内的第一道屏障。作为细胞壁主要成分之一,果胶在植物细胞壁累积重金属离子,缓解重金属毒害方面起关键性作用[5-6]。研究发现植物细胞壁中的果胶对Al3+、Cu2+、Cd2+以及其他金属离子的结合和富集起着关键性的作用[7-9]。一些新的研究发现细胞壁累积金属离子的多少与细胞壁果胶含量及果胶的甲酯化程度密切相关。果胶被分泌到胞外时甲基化程度很高,但果胶甲酯酶(Pectin methyl enzyme,PME)能使果胶去甲基化后转变为含有大量游离羧基的果胶酸[10]。含羧基果胶酸能够与重金属离子牢固结合,降低细胞质中游离态重金属离子浓度。
近年来,拟南芥(Arabidopsis thaliana)等模式植物细胞壁及果胶累积重金属的室内研究进展备受关注,但不同使用年限设施菜地重要果蔬作物番茄细胞壁果胶与Cd累积相关研究未见报道。本研究以不同年限设施菜地种植的常食果蔬番茄为材料,通过分析不同年限设施菜地土壤和番茄Cd累积量,以及细胞壁Cd累积量、果胶含量和PME活性等,研究细胞壁累积Cd含量与果胶含量以及PME活性的关系,以期进一步揭示设施菜地中番茄细胞壁果胶在番茄Cd累积过程中的作用机制。
1 材料与方法
1.1 实验材料
供试土壤和番茄植株采集于陕西省西安市郊区使用年限分别为2年和14年的设施菜地。每个年限分别布设了5个大棚,每个大棚按五点取样法采集0~20 cm土壤表层样品与对应番茄植株,最后将每个棚5个点的样品混合均匀,带回实验室备用。挑出土壤中的砂砾、植物根茎等杂物,过1 mm孔径筛。将过筛后的土壤样品装入自封袋,于4℃冰箱中保存备用。番茄植株用去离子水冲洗干净,吸水纸吸干水分,然后将茎和叶分开,分别放置在自封袋里,贴标签,于-80℃冰箱里保存备用。两种年限的土壤均为黄土母质,土壤类型为塿土,其基本理化性质见表1。
1.2 设施菜地土壤全Cd和有效Cd含量的测定
设施菜地土壤采用电热消解仪(DigiBlock ED54,LabTech)进行酸消解,取部分待测溶液用ICP-MS(i-CAP-Qc,ThermoFisher)进行全 Cd 含量测定[11]。土壤有效Cd含量依据国家标准GB/T 23739—2009方法进行测定[12]。
1.3 番茄植株Cd含量的测定
将茎、叶烘干,研磨。称取0.25 g待测样品于消解管内,加入10 mL HNO3,加盖摇匀,浸泡过夜。第二天放在电热消解仪(DigiBlock ED54,LabTech)中加热消解,其消解和样品Cd含量测定同1.2。
1.4 番茄细胞壁的提取和Cd含量的测定
番茄茎和叶细胞壁的提取参考Zhong等[13]的方法进行。将茎和叶分别加液氮磨碎,转移到50 mL离心管中,加入75%的冰乙醇浸没混匀,浸提3次,每次冰乙醇的用量为10 mL·g-1植物鲜重。混合物静置20 min后于4℃下5000×g离心10 min,去上清液后沉淀物再用 1∶7(鲜重/体积,g/mL)的冰丙酮、甲醇-三氯甲烷混合液(体积比1∶1)及甲醇溶液依次洗涤。每次洗涤,悬浮液静置10 min后离心。弃去上清液,沉淀冷冻干燥后,作为粗细胞壁,放入4℃冰箱保存备用。
番茄细胞壁消解和Cd含量测定同1.3。

表1 不同年限设施菜地土壤理化性质Table 1 Soil physical and chemical properties of greenhouse vegetable fields in different years
1.5 番茄细胞壁果胶的提取和Cd含量的测定
番茄细胞壁果胶的提取参考Zhong等[13]的方法进行。称取25 mg干燥的细胞样品,加入5 mL 0.5%的草酸铵缓冲液(含0.1%的NaHB4),沸水浴1 h,5000×g离心10 min,收集上清液,重复三次,上清液即为果胶。将上清液定容在25 mL容量瓶中,取部分溶液用ICP-MS进行Cd含量测定。
1.6 番茄细胞壁果胶糖醛酸含量的测定
果胶中糖醛酸含量的测定参考Willats等[14]、宾文等[15]方法测定。称取25 mg干燥的半乳糖醛酸,加水溶解,定容至25 mL容量瓶中,作为标准母液,样品冰浴10 min后,加入6 mL硫酸-硼酸盐溶液(四硼酸钠溶于浓硫酸),充分振荡,然后水浴加热5 min,立即放入冰水浴中冷却。加入0.1 mL的0.15%间苯二酚溶液(间苯二酚溶于0.5%的氢氧化钠溶液),空白组加入0.1 mL 0.5%氢氧化钠溶液,显色,充分振荡,静置20 min,于525 nm波长处测梯度溶液的吸光度,绘制标准曲线。对1.5节提取的果胶溶液进行上述操作,依据标准曲线计算样品中半乳糖醛酸含量。
1.7 果胶甲酯酶的提取和活性的测定
1.7.1 PME的提取
PME的提取参照Bordenave等[16]的方法。细胞壁经由NaCl溶液连续浸提,NaCl溶液的浓度梯度为0.2、0.4、0.5、1 mol·L-1细胞壁在 4 ℃条件下孵育 30 min,用巴氏玻璃吸管吸取上清液,上清液即为PME。
1.7.2 PME活性的测定
PME活性的测定参考Richarz等[17]的方法进行。分别将 80、120、160、200、240 μL PME 提取物加入到4 mL 底物溶液中[0.5%(m/V)柑橘果胶,0.2 mol·L-1NaCl,0.15%(m/V)甲基红,pH 6.8]。在 525 nm 处,用分光光度计测定溶液的吸光度值,绘制校准曲线。将8 μL PME提取物添加到4 mL底物溶液,在37℃孵育2 h,在525 nm处,用分光光度计测定溶液的吸光度值,得到相应的H+浓度,计算PME活性。
2 结果与分析
2.1 不同年限设施菜地土壤全Cd和有效Cd含量
不同年限设施菜地土壤全Cd和有效Cd含量如图1所示。2年和14年设施菜地土壤全Cd含量分别为 0.21、0.26 mg·kg-1,均未超过 0.3 mg·kg-1,符合土壤环境质量二级标准[18],但其含量是陕西省土壤Cd本底值的2.39、2.92倍(陕西省土壤Cd本底值为0.089 mg·kg-1)[19],且随着种植年限的增加,土壤中全 Cd 含量呈增加趋势。两个年限设施菜地土壤有效Cd含量存在显著差异(P<0.05),和2年的设施菜地相比,14年土壤有效Cd含量增加了195.53%。相对于全量,2年设施菜地中土壤有效Cd占23.46%,14年设施菜地中土壤有效Cd占53.51%。随着土壤全Cd含量的增加,有效Cd含量也随之增加,14年有效Cd含量是2年的2.96倍。
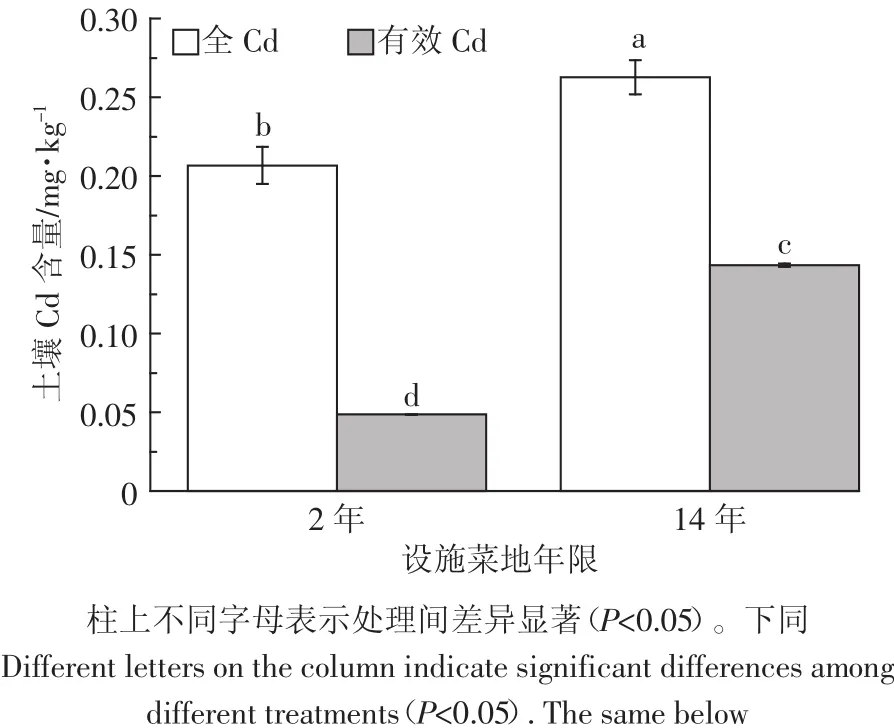
图1 不同年限设施菜地土壤全Cd和有效Cd含量Figure 1 Total and available Cd concentration in different planting years of greenhouse soil
2.2 不同年限设施菜地番茄Cd含量差异
不同年限设施菜地番茄植株Cd含量如图2所示。两个年限设施菜地番茄茎和叶片Cd累积量存在显著差异(P<0.05),和2年的设施菜地相比,14年番茄茎中Cd累积量增加了24.55%,叶片中Cd累积量增加了45.96%。在同年限中,番茄叶片中Cd累积量多于茎中的Cd累积量,在2年的设施菜地中,番茄叶Cd累积量是茎中的1.31倍,在14年的设施菜地中,番茄叶Cd累积量是茎中的1.54倍。

图2 不同年限设施菜地番茄植株Cd含量Figure 2 Cd concentration in tomato plant organs in different planting years of greenhouse soil
2.3 不同年限设施菜地对番茄细胞壁Cd含量的影响
从图3可以看出两个年限设施菜地番茄细胞壁Cd累积量存在显著差异(P<0.05),随着种植年限的增加,土壤Cd累积量增加,所种植的番茄细胞壁Cd含量有所上升。与2年的设施菜地番茄相比,14年番茄茎细胞壁Cd累积量增加了61.86%,叶片细胞壁Cd累积量增加了57.46%。同年限相比,在2年的设施菜地中,番茄叶片细胞壁Cd累积量是茎细胞壁Cd累积量的1.18倍,在14年的设施菜地中,番茄叶片细胞壁Cd累积量是茎细胞壁Cd累积量1.14倍。

图3 不同年限设施菜地番茄植株细胞壁Cd含量Figure 3 Cd concentration in tomato plant organs cell wall in different planting years of greenhouse soil
2.4 不同年限设施菜地对番茄细胞壁中果胶Cd含量的影响
不同年限设施菜地番茄细胞壁果胶Cd含量如图4所示。因设施菜地土壤累积Cd含量及番茄植株Cd含量有所不同,两个年限设施菜地番茄细胞壁果胶Cd累积量存在显著差异(P<0.05)。与2年的设施菜地番茄相比,14年番茄茎细胞壁果胶Cd累积量增加了725.15%,叶片细胞壁果胶Cd累积量增加了522.20%。同年限相比,在2年的设施菜地中,番茄叶片果胶Cd累积量是茎果胶Cd累积量的1.61倍,在14年的设施菜地中,番茄叶片果胶Cd累积量是茎果胶Cd累积量的1.21倍。

图4 不同年限设施菜地番茄细胞壁果胶Cd含量Figure 4 Cd concentration in cell wall pectin in different planting years of greenhouse soils
2.5 不同年限设施菜地果胶半乳糖醛酸含量的差异
双子叶植物细胞壁的果胶中半乳糖醛酸的含量比较稳定,一般来说,半乳糖醛酸的含量可以用来衡量果胶含量水平[20]。不同年限设施菜地番茄细胞壁果胶半乳糖醛酸含量如图5所示,种植年限越长,土壤Cd累积量越多的菜地,其细胞壁果胶半乳糖醛酸含量越多。在两种年限的设施菜地中,番茄叶片和茎细胞壁果胶半乳糖醛酸含量都存在显著差异(P<0.05)。和2年相比,14年番茄茎中果胶半乳糖醛酸含量提高了104.51%,叶片提高了127.45%。同年限相比,在2年的设施菜地中,番茄叶细胞壁果胶半乳糖醛酸的含量是茎中的1.88倍,在14年的设施菜地中,番茄叶细胞壁果胶半乳糖醛酸的含量是茎中的2.09倍。
研究发现细胞壁Cd累积量和果胶半乳糖醛酸含量具有很好的线性相关关系(R2=0.670 6),随着细胞壁Cd累积量的增多,果胶半乳糖醛酸的含量也随之增多,由此可推测番茄果胶含量在果胶累积重金属Cd中起一定的作用。
2.6 不同年限设施菜地果胶甲酯酶活性的差异
不同年限设施菜地番茄细胞壁PME活性(nmol H+·mg-1·h-1)如图 6 所示,种植年限越长,其番茄细胞壁PME活性越高。在两种年限的设施菜地中,番茄叶片和茎细胞壁PME活性存在差异。和2年相比,14年番茄茎中PME活性提高了10.56%,叶片果胶PME活性提高了9.39%。同年限相比,在2年的设施菜地中,番茄茎细胞壁PME活性是叶的1.07倍,在14年的设施菜地中,番茄叶细胞壁PME活性是茎中的1.06倍。

图5 不同年限设施菜地番茄细胞壁果胶半乳糖醛酸含量Figure 5 Galacturonic acid contention in the cell wall in different planting years of greenhouse soils
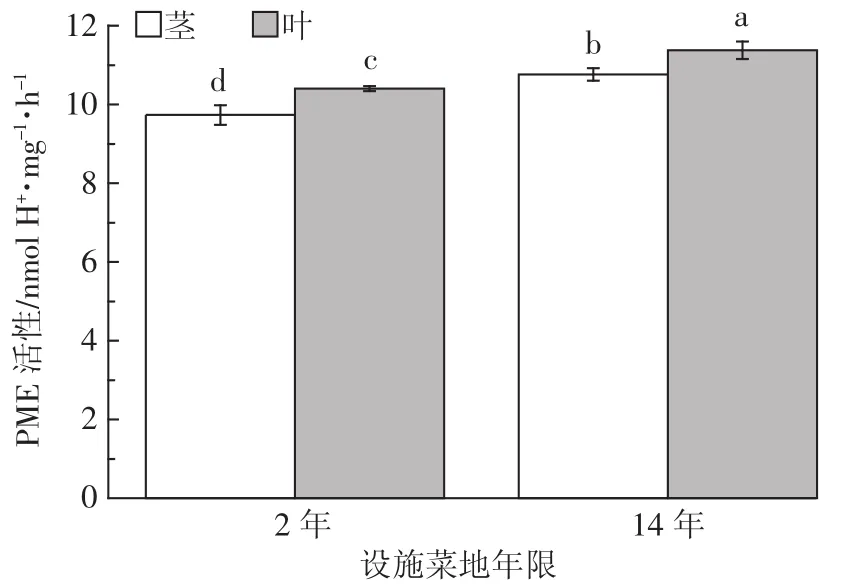
图6 不同年限设施菜地番茄细胞壁PME活性Figure 6 PME activity in the cell wall in different planting years of greenhouse soils
研究发现细胞壁Cd累积量和PME活性具有很好的线性相关关系(R2=0.891),随着细胞壁Cd累积量的增多,PME的活性也随之升高,由此可推测番茄PME在果胶累积重金属Cd中起一定的作用。
3 讨论
近年来,设施菜地重金属的累积及其安全风险问题正逐步引起社会各界高度关注。设施菜地常年处于半封闭状态,复种指数高,无雨水沉降,使得因污水灌溉、化肥施用等带来的重金属不断累积。井永苹等[21]以山东省寿光市设施蔬菜种植基地为对象,探讨了不同种植年限对设施菜地土壤重金属累积量的影响,结果显示土壤Cd含量随种植年限增加而升高。本文的研究结果与其一致,如图1所示,14年的设施菜地土壤全Cd含量是2年的1.27倍,有效Cd含量是2年的2.96倍。随着土壤中Cd累积量的增加,番茄茎和叶Cd的含量均呈明显的增加趋势。植物根系从土壤中吸收重金属,向上运输至茎和叶等器官,由于转移能力和转运机制的不同,植物的各器官对重金属的吸收、累积能力往往存在较大差异,在许多超富集植物中,叶片对重金属的富集能力高于茎[22-24]。本研究中,番茄叶片中Cd累积量明显多于茎中Cd累积量。
研究表明细胞壁是响应重金属胁迫的功能信号分子和代谢所在的位点,参与植物对重金属胁迫的响应过程[25-26]。在重金属胁迫环境中,植物细胞壁可富集大量的金属离子,阻碍其进入原生质体影响叶绿体等细胞器的活性。如茶树根细胞壁可累积大量的铅离子[27]。如图2所示,番茄叶片和茎细胞壁均可累积Cd2+,且设施菜地种植年限越长,细胞壁累积Cd含量越多。
细胞壁的各种组分尤其是多糖在固定重金属方面发挥重要的作用。果胶是植物细胞壁主要组成成分之一,富含多种带负电荷的有机官能团,如羟基、羧基、氨基、醛基、巯基等,可提供重金属离子结合位点,它们对重金属离子的吸附络合发挥关键性作用[28]。本研究发现番茄细胞壁果胶累积Cd含量占了细胞壁Cd总含量的4.36%~22.25%。王梦等[27]研究发现茶树根细胞壁吸附Pb过程中,20%的Pb是吸附在果胶上的;徐劼等[29]对芹菜细胞壁进行果胶酶改性处理,降低果胶含量后,其对Cd的累积量相对降低了40.5%。果胶半乳糖醛酸含量和细胞壁Cd累积量显著正相关,说明番茄果胶在细胞壁Cd累积中起重要作用。果胶甲酯酶促使甲酯化果胶上的甲醇基从多聚半乳糖醛酸羧基上脱离,降低果胶甲酯化程度。Krzes覥owska[30]研究发现高浓度的铜刺激细胞内产生果胶甲酯酶,催化甲醇基发生酯化反应形成羧基,从而具有更多的铜离子的结合位点,使之能结合更多的铜。李学文等[31]指出根尖细胞壁PME活性越高,果胶甲基酯化程度越低。PME活性和细胞壁Cd累积量的线性关系说明番茄细胞壁果胶PME在Cd累积过程中的重要作用。随着种植年限的增加,植物体内Cd累积量的增多,诱导PME活性增加。由此可见,PME直接或间接地对植物重金属耐性机制起作用,反之重金属也会影响PME的活性。
4 结论
(1)随种植年限的增加,番茄植株各器官Cd累积量增加。此外,不同器官中Cd累积量存在差异,Cd在番茄叶片、叶片细胞壁以及叶片细胞壁果胶的累积量均大于茎部。
(2)随种植年限的增加,番茄细胞壁Cd累积量增加。14年设施菜地的番茄细胞壁Cd含量显著高于2年。同时,14年设施菜地的番茄细胞壁果胶Cd含量大于2年,且果胶含量和细胞壁Cd累积量呈显著正相关。
(3)随种植年限的增加,番茄细胞壁PME活性增加。14年设施菜地的番茄细胞壁PME活性大于2年,且PME活性和细胞壁Cd累积量呈显著正相关。
[1]中华人民共和国环境保护部,国土资源部.全国土壤污染状况调查公报[R].北京:中华人民共和国环境保护部,国土资源部,2014:1-5.Ministry of Environmental Protection of PRC,Ministry of Land and Resources of PRC.National investigation report on soil pollution[R].Beijing:Ministry of Environmental Protection of PRC,Ministry of Land and Resources of PRC,2014:1-5.
[2]李 婧,周艳文,陈 森,等.我国土壤镉污染现状、危害及其治理方法综述[J].安徽农学通报,2015,21(24):104-107.LI Jing,ZHOU Yan-wen,CHEN Sen,et al.Actualities,damage and management of soil cadmium pollution in China[J].Anhui Agricultural Science Bulletin,2015,21(24):104-107.
[3]Zhang X,Gao B,Xia H.Effect of cadmium on growth,photosynthesis,mineral nutrition and metal accumulation of bana grass and vetiver grass[J].Ecotoxicology&Environmental Safety,2014,106:102-108.
[4]Lópezmillán A F,Sagardoy R,Solanas M,et al.Cadmium toxicity in tomato(Lycopersicon esculentum)plants grown in hydroponics[J].Environmental&Experimental Botany,2009,65(3):376-385.
[5]潘伟槐,郑仲仲,郭天荣,等.果胶甲基酯酶PME参与调控大麦根尖铝毒敏感性[J].浙江大学学报(理学版),2011,38(3):326-332.PAN Wei-huai,ZHENG Zhong-zhong,GUO Tian-rong,et al.Involvement of PME activity in regulation of Al toxic sensitivity in the root tips of barley[J].Journal of Zhejiang University(Science Edition),2011,38(3):326-332.
[6]Dalcorso G,Farinati S,Furini A.Regulatory networks of cadmium stress in plants[J].Plant Signaling&Behavior,2010,5(6):663-667.
[7]Muschitz A,Riou C,Mollet J C,et al.Modifications of cell wall pectin in tomato cell suspension in response to cadmium and zinc[J].Acta Physiologiae Plantarum,2015,37(11):245.
[8]Douchiche O,Rihouey C,Schaumann A,et al.Cadmium-induced alterations of the structural features of pectins in flax hypocotyl[J].Planta,2007,225(5):1301-1312.
[9]Colzi I,Doumett S,Bubba M D,et al.On the role of the cell wall in the phenomenon of copper tolerance in Silene paradoxa L.[J].Environmental&Experimental Botany,2011,72(1):77-83.
[10]Micheli F.Pectin methylesterases:Cell wall enzymes with important roles in plant physiology[J].Trends in Plant Science,2001,6(9):414-419.
[11]董明芳,郭军康,冯人伟,等.Fe2+和Mn2+对水稻根表铁膜及镉吸收转运的影响[J].环境污染与防治,2017,39(3):249-253.DONG Ming-fang,GUO Jun-kang,FENG Ren-wei,et al.Effects of Fe2+and Mn2+on rice root iron plaque formation and Cd uptake and transportation[J].Environmental Pollution&Control,2017,39(3):249-253.
[12]国家质量监督检验检疫总局,中国国家标准化管理委员会.GB/T 23739—2009土壤质量有效态铅和镉的测定原子吸收方法[S].北京:中国标准出版社,2009.General Administration of Quality Supervision,Inspection and Quarantine of the People′s Republic of China,Standardization Administration of China.GB/T 23739—2009 Soil quality-Analysis of available lead and cadmium in soils-Atomic absorption spectrometry[S].Beijing:Standards Press of China,2009.
[13]Zhong H,L覿uchli A.Changes of cell wall composition and polymer size in primary roots of cotton seedlings under high salinity[J].Journal of Experimental Botany,1993,44(261):773-778.
[14]Willats W G,Knox J P,Mikkelsen J D.Pectin:New insights into an old polymer are starting to gel[J].Trends in Food Science&Technology,2006,17(3):97-104.
[15]宾 文,于荣敏,白秀峰.人工培养蛹虫草多糖的研究[J].沈阳药科大学学报,2000,17(5):361-364.BIN Wen,YU Rong-min,BAI Xiu-feng.Studies on polysaccharide in the tissue cultures of cordyceps militaris[J].Journal of Shenyang Pharmaceutical University,2000,17(5):361-364.
[16]Bordenave M,Goldberg R.Immobilized and free apoplastic pectinmethylesterases in mung bean hypocotyl[J].Plant Physiology,1994,106(3):1151-1156.
[17]Richard L,Qin L X,Gadal P,et al.Molecular cloning and characterisation of a putative pectin methylesterase cDNA in Arabidopsis thaliana(L.)[J].Febs Letters,1994,355(2):135-139.
[18]环境保护部,国家质量监督检验检疫总局.GB 15618—2008土壤环境质量标准(修订)[S].北京:中国标准出版社,2008.Ministry of Environmental Protection of PRC,General Administration of Quality Supervision,Inspection and Quarantine of the People′s Republic of China.GB 15618—2008 Environmental quality standard for soils[S].Beijing:Standards Press of China,2008.
[19]中国环境监测总站.中国土壤元素背景值[M].北京:中国环境科学出版社,1990.China National Environmental Monitoring Centre.Background values of soil elements in China[M].Beijing:China Environmental Science Press,1990.
[20]Caffall K H,Mohnen D.The structure,function,and biosynthesis of plant cell wall pectic polysaccharides[J].Carbohydrate Research,2009,344(14):1879-1900.
[21]井永苹,李 彦,薄录吉,等.不同种植年限设施菜地土壤养分、重金属含量变化及主导污染因子解析[J].山东农业科学,2016,48(4):66-71.JING Yong-ping,LI Yan,BO Lu-ji,et al.Variation of soil nutrient and heavy metal accumulation in greenhouse soil with cultivation years and analysis on main pollution factor[J].Shandong Agricultural Sciences,2016,48(4):66-71.
[22]Seo K W,Son Y,Rhoades C C,et al.Seedling growth and heavy metal accumulation of candidate woody species for revegetating Korean mine spoils[J].Restoration Ecology,2008,16(4):702-712.
[23]茹淑华,张国印,贡冬梅,等.铜、锌、铅和镉复合污染对番茄生长和重金属累积规律的影响[J].华北农学报,2013,28(增刊1):371-375.RU Shu-hua,ZHANG Guo-yin,GONG Dong-mei,et al.Effects of combined pollution of Cu,Zn,Pb and Cd on growth of tomato and heavy metal accumulation regularity[J].Acta Agriculturae Boreali-Sinica,2013,28(Suppl 1):371-375.
[24]Deng H,Li M S,Chen Y X,et al.Accumulation characteristics of Cd in Polygonum pubescens Bl[J].Agricultural Science&Technology,2010,3(8):580-586.
[25]Chen G C,Liu Y Q,Wang R M,et al.Cadmium adsorption by willow root:The role of cell walls and their subfractions[J].Environmental Science and Pollution Research,2013,20(8):5665-5672.
[26]Dalcorso G,Farinati S,Furini A.Regulatory networks of cadmium stress in plants[J].Plant Signaling&Behavior,2010,5(5):663-667.
[27]王 梦,段德超,徐 辰,等.茶树根细胞壁不同组分对铅的吸附性能及其功能团的傅里叶红外光谱学研究[J].生态学报,2015,35(6):1743-1751.WANG Meng,DUAN De-chao,XU Chen,et al.Adsorption ability of cell wall(CW)components in roots of tea plant(Camellia sinensis L.)to Pb and FTIR spectra of their functional groups[J].Acta Ecologica Sinica,2015,35(6):1743-1751.
[28]Haynes R J.Ion exchange properties of roots and ionic interactions within the root apoplasm:Their role in ion accumulation by plants[J].The Botanical Review,1980,46(1):75-99.
[29]徐 劼,保积庆.芹菜根细胞壁对镉的吸附固定机制及其FTIR表征研究[J].环境科学学报,2015,35(8):2605-2612.XU Jie,BAO Ji-qing.Adsorption and fixation mechanism of cadmium on celery(Apium graveolens L.)root cell wall and the analysis of FTIR spectra[J].Acta Scientiae Circumstantiae,2015,35(8):2605-2612.
[30]Krzes覥owska M.The cell wall in plant cell response to trace metals:Polysaccharideremodelinganditsroleindefensestrategy[J].Acta Physiologiae Plantarum,2011,33(1):35-51.
[31]李学文,李亚林,杨 锦,等.豌豆不同耐铝品种根尖细胞壁果胶及其甲基酯化度的差异[J].植物营养与肥料学报,2016,22(3):729-735.LI Xue-wen,LI Ya-lin,YANG Jin,et al.Dimrences of pectin content and methylation in root cell wall of pea(Pisum satvium)cultivars with different Al tolerance[J].Journal of Plant Nutrition and Fertilizer,2016,22(3):729-735.
猜你喜欢
数学小灵通·3-4年级(2022年5期)2022-12-31
数学小灵通·3-4年级(2021年5期)2021-07-16
烟台大学学报(自然科学与工程版)(2021年1期)2021-03-19
三农资讯半月报(2020年18期)2020-10-14
读友·少年文学(清雅版)(2020年10期)2020-03-19
合肥师范学院学报(2020年6期)2020-03-10
酿酒科技(2019年1期)2019-02-25
数码世界(2018年1期)2018-12-23
浙江工业大学学报(2017年5期)2018-01-22
中国果菜(2017年8期)2017-01-14
