
肾素/肾素前体受体在高血压中的进展
2018-03-21 07:54顾婕综述姜一农杨晓蕾审校
心血管病学进展 2018年1期
顾婕 综述 姜一农 杨晓蕾 审校
(1.大连医科大学,辽宁 大连 116000; 2.大连医科大学附属第一医院 心血管病医院,辽宁 大连 116000)
2002年,Nguyen等[1]报道并克隆出一种膜表面蛋白分子——肾素/肾素前体受体[(pro)renin receptor,(P)RR],可与肾素及肾素前体结合,增强组织肾素-血管紧张素系统(renin-angiotensin-system,RAS)活性,同时诱导非RAS的细胞内信号转导,如在丝裂原激活蛋白激酶(mitogen-activated protein kinases,MAPK)通路中引起细胞增殖,上调促转化生长因子-β(transforming growth factor-β,TGF-β)和纤维化因子等表达增加。随后研究发现(P)RR的功能与V-ATPase有重要联系,且(P)RR是Wnt通路必不可少的成员之一,与体内酸碱平衡的调节、自噬降解及细胞增殖发育密切相关。(P)RR几乎表达于体内所有的组织中,参与多种疾病的发生发展。无论是在血管紧张素Ⅱ(angiotensinⅡ,AngⅡ)依赖或非依赖的途经中,(P)RR信号通路都与神经源性高血压、心肌病、肾小球硬化、脉络膜新生血管等疾病的发生相关。
1 (P)RR的结构
(P)RR是由350个氨基酸组成的相对分子质量为35 000~39 000的跨膜蛋白,包括四个结构域:与肾素和肾素前体结合的氨基末端、构成胞外结构的羧基末端,以及单次跨膜区和胞内短二聚体尾。肾素/肾素前体与(P)RR结合后,会诱导其分子构象发生变化,使抑制性前肽片段水解,暴露出活性部位,从而激活肾素(图1)。
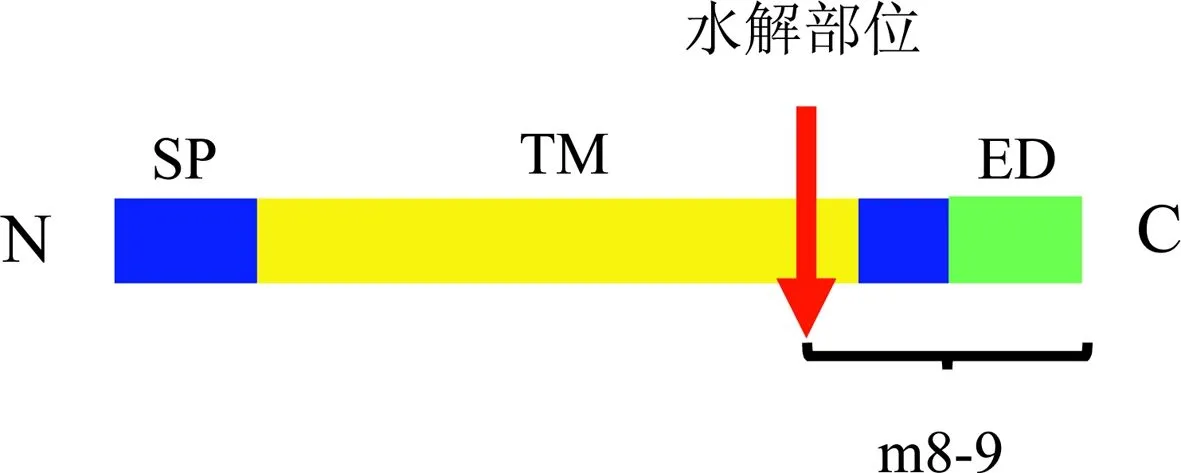
a

b
注:a图(P)RR的结构域和其水解片段,红色箭头为水解部位。N:氨基末端;C:羧基末端;SP:信号肽;TM:跨膜结构域;ED:胞外结构域;m8-9:V-ATPase的辅助蛋白。b图(P) RR插入多功能的V-ATPase中。
图1 (P)RR的基本结构
(P)RR可被水解,其截断的跨膜区和在细胞质内的部分被认为是空泡型H+-ATPase的一种相对分子质量8 900的辅助蛋白(m8-9)(图1)。此外,Cousin 等在几种不同的细胞培养基中发现一种相对分子质量28 000的可溶性肾素/肾素前体受体[soluble (pro)renin receptor,s(P)RR],可由蛋白酶(如丝氨酸蛋白酶和金属蛋白酶)切割全长(P)RR产生。
2 (P)RR的功能
2.1 (P)RR与MAPK通路
(P)RR由ATP6AP2基因编码,定位于人类,猴子(黑猩猩、猕猴),啮齿类动物(小鼠、大鼠)和其他哺乳动物(如狗、牛)的X染色体,广泛分布于心、脑、肾、肝、胎盘和胰腺等组织,但是其生理作用尚未完全阐明[1]。(P)RR的基因序列表明它拥有多重身份:H+载体、溶酶体辅助蛋白2、ATP酶、内质网I型跨膜接头前体。 因此,肾素/肾素前体与(P)RR的结合可以完全独立于RAS,诱导信号产生。事实上,在肾小球系膜细胞、集合管细胞、血管平滑肌细胞,以及单核细胞和神经细胞上,肾素/肾素前体与(P)RR结合会激活MAPK信号通路,增强细胞外信号调节激酶(extracellular signal-regulated kinase 1/2,ERK 1/2)的磷酸化,引起细胞增殖,增加TGF-β表达量,上调纤维化因子的表达水平,比如纤溶酶原激活物抑制剂(plasminogen activator inhibitor,PAI),以及胶原蛋白和纤维连接蛋白等(图2)。此外,在血管平滑肌细胞中,肾素/肾素前体与(P)RR结合会激活丝氨酸/苏氨酸激酶;在心肌细胞中,会激活p38 MAPKs和热休克蛋白27(Hsp27),并调节肌动蛋白细胞骨架动力学。
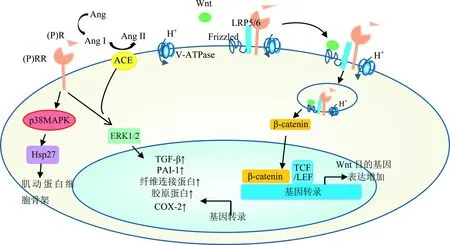
注:Wnt 与Frizzled、LRP5/6的复合物(Fz/LRP5/6)结合,使β-catenin的磷酸化被抑制,β-catenin进入细胞核后与T细胞因子/淋巴细胞增强子复合物(T-cell factor/lymphocyte enhancer binding factor, TCF/LEF)结合,激活一系列基因转录。Fz/LRP5/6形成和其在运输囊泡内磷酸化是Wnt/β-catenin 经典途径的关键过程。(P)RR和V-ATPase与其运输囊泡的磷酸化密切相关。ACE:血管紧张素转化酶; COX:环氧合酶;(P)R:肾素受体。
图2 (P)RR的功能和信号传导
2.2 (P)RR与V-ATPase
另有研究表明(P)RR 对V-ATPase的完整性具有非常重要的作用。V-ATPase是多亚基蛋白质,由V0质子转位结构域和V1泵结构域,以及2种相关的蛋白:Ac45 和(P)RR构成(图1)。V-ATPase几乎存在于所有类型的细胞中,主要在细胞内的质膜上,对膜泡的运输、蛋白降解及偶联转运具有重要作用。在一些细胞中,V-ATPase也大量存在于细胞膜上,例如,在集合管闰细胞膜上,V-ATPase用于调节全身酸碱平衡。另外,敲除(P)RR基因的心肌细胞和足细胞都呈现高度的空泡化并发生有害的自噬性降解[2],而这种自噬缺陷则是由于V0亚基选择性下调细胞内囊泡脱氧造成[3]。敲除(P)RR基因的血管平滑肌细胞[4],感光细胞[5]和输尿管芽[6]也会出现缺乏V-ATPase功能相关的表型。因此,(P)RR对V-ATPase的结构、功能、稳定都是必不可少的。
2.3 (P)RR与经典Wnt/β-catenin信号通路
Wnt/β-catenin信号依赖于V-ATPase介导的磷酸化信号转导机制,而(P)RR在其中起着不可或缺的作用[7],由于后者不具备RAS活性,因此可证实(P)RR有独立于肾素/肾素前体的功能。Wnts是分泌性糖蛋白家族,经典的Wnt信号已被证实对细胞生长发育非常重要。Cruciat等证实了(P)RR的消融会破坏V-ATPase完整性,而无法形成质子梯度,阻碍Wnt信号转导的磷酸化(图2)。在过表达(P)RR基因的大鼠模型中,不仅可见肾素/肾素前体与(P)RR结合的效应,同时也可见与Wnt/β-catenin经典信号通路相关的效应[8]。在糖尿病患者及其动物模型中,均可见肾小球和足细胞的Wnt/β-catenin信号增强[9],启动下游TGF-β的表达增强,诱导包括纤溶酶原活化物抑制剂在内的促纤维化因子的产生[10]。全(P)RR基因敲除的小鼠在胚胎早期即死亡,这可能与阻断Wnt信号通路有关,因为Wnt信号通路是小鼠早期胚胎发育的必要条件,而这给(P)RR的研究带来了许多困难。
3 (P)RR与高血压
3.1 (P)RR与神经源性高血压
研究指出大脑是(P)RR表达的重要部位[11]。在控制全身电解质平衡和调节心血管的功能方面,大脑中的RAS起着重要的作用,脑内的AngⅡ被证实能够增强交感神经兴奋活性,造成血压升高、口渴等心血管功能紊乱和电解质失衡[8],尤其是会导致神经源性高血压形成[12-13];但是大脑中的肾素水平及酶活性很低,不足以介导AngⅡ的形成并引起其下游的影响,而循环中的肾素也不能通过血脑屏障,这就说明AngⅡ是在大脑局部内形成。研究表明,在大脑中广泛表达的(P)RR[14]可能是脑内AngⅡ的主要来源[15]。因此,在中枢神经系统中,(P)RR主要通过AngⅡ的形成调节血压,诱导神经源性高血压产生。
大脑中(P)RR的激活会刺激交感神经和下丘脑室旁核神经元放电活动增加。(P)RR主要集中在垂体前叶,在室旁核和视上核与加压素和催产素共定位,通过中枢控制水电解质代谢和血压。敲除自发性高血压大鼠视上核的(P)RR,能改善年龄依赖性高血压,降低心率和血管加压素的释放。特异性敲除神经元(P)RR基因可以预防醋酸脱氧皮质酮-盐敏感性高血压[13,16]。在醋酸脱氧皮质酮-盐敏感性高血压模型中,模型组小鼠脑AngⅡ增加约是野生型的5倍,而特异性敲除神经元(P)RR基因的模型小鼠中则不存在AngⅡ增加[16]。将肾素原前段20个氨基酸多肽作为(P)RR的拮抗剂注入脑内,可以观察到与氯沙坦相似的降压效果[17];但是其血浆AngⅡ水平仍是正常野生型小鼠的3~4倍,大脑皮层和下丘脑中AngⅡ也仅降低了约25%。因此,PRO20作为(P)RR的拮抗剂产生的降压效应,并不只是通过抑制RAS产生,还可能通过MAPK通路和Wnt通路等非AngⅡ依赖途径产生降压作用[17]。并且,在中枢神经系统中,(P)RR对血压的调控作用不仅仅是通过RAS产生。
3.2 肾脏(P)RR与血压的调节
在肾脏中,(P)RR定位于肾小管、肾小球系膜细胞、足细胞、远曲小管及集合管等,主要参与肾脏的发育[7,18]、尿液浓缩[19-20]、水钠代谢及血压调节[21]。研究表明,集合管(P)RR的表达水平在肾脏中最高,大鼠经AngⅡ刺激后引起肾脏(P)RR的基因转录增加。而在AngⅡ非依赖途径中,集合管(P)RR与肾素/肾素前体结合激活ERK 1/2、MAPK、PI3K等途径。非选择性过表达人(P)RR的转基因大鼠,表现为正常血压但存在进展性肾病和蛋白尿[22],选择性动脉平滑肌(P)RR过表达的转基因大鼠,表现出高血压和心动过速,但是未出现肾脏选择肾脏表型的改变。也有研究表明,(P)RR通过集合管的上皮钠通道蛋白参与Ang Ⅱ 诱导的高血压调控[23-24]。这些研究说明肾脏(P)RR参与血压与肾功能的调节[25]。
3.3 其他
脂肪细胞 (P)RR缺陷的小鼠,可以检测到其血浆中的s(P)RR[26]和血压[27]明显增加。胎盘脂肪细胞(P)RR的过表达,以及孕妇血浆s(P)RR的水平升高均与妊娠期血压升高相关[27]。在视网膜血管内皮细胞上,肾素/肾素前体刺激(P)RR上调促进血管生成和炎症反应[28]。因此,(P)RR在许多组织中都参与血压的调控。
4 (P)RR与其他心血管疾病
氧化型低密度脂蛋白刺激人主动脉平滑肌细胞可以引起(P)RR表达增加,说明(P)RR在动脉粥样硬化中具有一定作用[29]。心房颤动患者血浆中(P)RR显著增加,而小鼠心脏过表达(P)RR基因会诱导心房颤动产生,心房特异性表达(P)RR基因则出现异常的心房电生理、心房肥大及纤维化,最终诱导心房颤动产生[30]。
5 (P)RR的阻滞剂
除了前段20个氨基酸多肽可以作为(P)RR的拮抗剂外,柄区肽(HRP)也曾被认为是(P)RR的一种拮抗剂;但是,很多文章都无法证明HRP可以阻断肾素(前体)诱导的信号转导通路。甚至有人认为HRP是(P)RR的部分激动剂,还有人认为与HRP结合的细胞并不表达(P)RR。因此,对 (P)RR阻滞剂的使用尚存在争议。
6 结论
过去十几年对(P)RR的研究显示了其与RAS的密切关系,同时也证实了(P)RR的存在完全独立于RAS的作用。它可能参与多个受体的信号转导通路,在各种心血管疾病,尤其是高血压等疾病中起着关键作用。虽然(P)RR拮抗剂的作用尚不能得到完全的证实,但是对(P)RR拮抗剂的研究对许多疾病的发生发展及治疗都提供了新的思路及靶点。
[1] Nguyen G,Delarue F,Burckle C,et al.Pivotal role of the renin/prorenin receptor in angiotensin Ⅱ production and cellular responses to renin[J].J Clin Invest,2002,109(11):1417-1427.
[2] Kissing S,Hermsen C,Repnik U,et al.Vacuolar ATPase in phagosome-lysosome fusion[J].J Biol Chem,2015,290(22):14166-14180.
[3] Oshima Y,Kinouchi K,Ichihara A,et al.Prorenin receptor is essential for normal podocyte structure and function[J].J Am Soc Nephrol,2011,22(12):2203-2212.
[4] Kurauchi-Mito A,Ichihara A,Bokuda K,et al.Significant roles of the (pro)renin receptor in integrity of vascular smooth muscle cells[J].Hypertens Res,2014,37(9):830-835.
[5] Kanda A,Noda K,Yuki K,et al.Atp6ap2/(pro)renin receptor interacts with Par3 as a cell polarity determinant required for laminar formation during retinal development in mice[J].J Neurosci,2013,33(49):19341-19351.
[6] Song R,Preston G,Ichihara A,et al.Deletion of the prorenin receptor from the ureteric bud causes renal hypodysplasia[J].PLoS One,2013,8(5):e63835.
[7] Bernhard SM,Seidel K,Schmitz J,et al.The (pro)renin receptor ((P)RR) can act as a repressor of Wnt signalling[J].Biochem Pharmacol,2012,84(12):1643-1650.
[8] Song R,Janssen A,Li Y,et al.Prorenin receptor controls renal branching morphogenesis via Wnt/beta-catenin signaling[J].Am J Physiol Renal Physiol,2017,312(3):F407-F417.
[9] Kato H,Gruenwald A,Suh JH,et al.Wnt/beta-catenin pathway in podocytes integrates cell adhesion,differentiation,and survival[J].J Biol Chem,2011,286(29):26003-26015.
[10] He W,Tan R,Dai C,et al.Plasminogen activator inhibitor-1 is a transcriptional target of the canonical pathway of Wnt/beta-catenin signaling[J].J Biol Chem,2010,285(32):24665-24675.
[11] Brugts JJ,Isaacs A,de Maat MP,et al.A pharmacogenetic analysis of determinants of hypertension and blood pressure response to angiotensin-converting enzyme inhibitor therapy in patients with vascular disease and healthy individuals[J].J Hypertens,2011,29(3):509-519.
[12] Grobe JL,Grobe CL,Beltz TG,et al.The brain renin-angiotensin system controls divergent efferent mechanisms to regulate fluid and energy balance[J].Cell Metab,2010,12(5):431-442.
[13] Li W,Peng H,Mehaffey EP,et al.Neuron-specific (pro)renin receptor knockout prevents the development of salt-sensitive hypertension[J].Hypertension,2014,63(2):316-323.
[14] Narumi K,Hirose T,Sato E,et al.A functional (pro)renin receptor is expressed in human lymphocytes and monocytes[J].Am J Physiol Renal Physiol,2015,308(5):F487-499.
[15] Takahashi K,Hiraishi K,Hirose T,et al.Expression of (pro)renin receptor in the human brain and pituitary,and co-localisation with arginine vasopressin and oxytocin in the hypothalamus[J].J Neuroendocrinol,2010,22(5):453-459.
[16] Zubcevic J,Jun JY,Lamont G,et al.Nucleus of the solitary tract (pro)renin receptor-mediated antihypertensive effect involves nuclear factor-kappaB-cytokine signaling in the spontaneously hypertensive rat[J].Hypertension,2013,61(3):622-627.
[17] Li W,Sullivan MN,Zhang S,et al.Intracerebroventricular infusion of the (pro)renin receptor antagonist PRO20 attenuates deoxycorticosterone acetate-salt-induced hypertension[J].Hypertension,2015,65(2):352-361.
[18] Yosypiv IV.Prorenin receptor in kidney development[J].Pediatr Nephrol,2017,32(3):383-392.
[19] Jensen BL.Prorenin receptor,a necessary component in urine concentration mechanism[J].J Am Soc Nephrol,2016,27(10):2919-2921.
[20] Ramkumar N,Stuart D,Calquin M,et al.Nephron-specific deletion of the prorenin receptor causes a urine concentration defect[J].Am J Physiol Renal Physiol,2015,309(1):F48-56.
[21] Quadri S,Siragy HM.(Pro)renin receptor contributes to regulation of renal epithelial sodium channel[J].J Hypertens,2016,34(3):486-494;discussion 494.
[22] Kaneshiro Y,Ichihara A,Sakoda M,et al.Slowly progressive,angiotensin Ⅱ-independent glomerulosclerosis in human (pro)renin receptor-transgenic rats[J].J Am Soc Nephrol,2007,18(6):1789-1795.
[23] Peng K,Lu X,Wang F,et al.Collecting duct (pro)renin receptor targets ENaC to mediate angiotensin Ⅱ-induced hypertension[J].Am J Physiol Renal Physiol,2017,312(2):F245-F253.
[24] Ramkumar N,Stuart D,Mironova E,et al.Renal tubular epithelial cell prorenin receptor regulates blood pressure and sodium transport[J].Am J Physiol Renal Physiol,2016,311(1):F186-194.
[25] Burckle CA,Jan Danser AH,Muller DN,et al.Elevated blood pressure and heart rate in human renin receptor transgenic rats[J].Hypertension,2006,47(3):552-556.
[26] Tan P,Shamansurova Z,Bisotto S,et al.Impact of the prorenin/renin receptor on the development of obesity and associated cardiometabolic risk factors[J].Obesity(Silver Spring),2014,22(10):2201-2209.
[27] Wu CH,Mohammadmoradi S,Thompson J,et al.Adipocyte (pro)renin-receptor deficiency induces lipodystrophy,liver steatosis and increases blood pressure in male mice[J].Hypertension,2016,68(1):213-219.
[28] Zhu T,Miller AG,Deliyanti D,et al.Prorenin stimulates a pro-angiogenic and pro-inflammatory response in retinal endothelial cells and an M1 phenotype in retinal microglia[J].Clin Exp Pharmacol Physiol,2015,42(5):537-548.
[29] Cheng Y,Liu X,Zhang L,et al.Effect of oxidized low-density lipoprotein on the expression of the prorenin receptor in human aortic smooth muscle cells[J].Mol Med Rep,2015,11(6):4341-4344.
[30] Lian H,Wang X,Wang J,et al.Heart-specific overexpression of (pro)renin receptor induces atrial fibrillation in mice[J].Int J Cardiol,2015,184:28-35.
猜你喜欢
湖北农业科学(2022年11期)2022-07-18
天津医科大学学报(2021年1期)2021-12-05
世界睡眠医学杂志(2021年4期)2021-07-03
实用肿瘤学杂志(2020年4期)2020-12-08
中华养生保健(2020年2期)2020-11-16
世界科学技术-中医药现代化(2020年2期)2020-07-25
牡丹江医学院学报(2020年2期)2020-07-01
天津医科大学学报(2019年3期)2019-08-13
中国粮油学报(2019年4期)2019-07-12
中国老年学杂志(2015年9期)2015-01-31
