
花生四烯酸对TNF-α/放线菌素D诱导内皮细胞凋亡的影响及作用机制
2018-04-20 06:40陈积雄黄晓燕林文婷何扬利符秀虹曾敏汪道文
山东医药 2018年12期
陈积雄,黄晓燕,林文婷,何扬利,符秀虹,曾敏,汪道文
(1 海南省人民医院,海口570030;2华中科技大学附属同济医院)
内皮细胞凋亡在心血管疾病的发生及发展过程中起重要作用,是冠心病、心力衰竭等疾病的始动因素[1]。花生四烯酸经表氧化酶代谢可生成多种环氧二十碳四烯酸(EET),对维持心血管系统稳定至关重要[2,3]。研究证实,TNF-α可促进内皮细胞凋亡;EET可抑制TNF-α诱导的内皮细胞凋亡,但具体作用机制尚未阐明[4,5]。2016年5月~2017年5月,本研究观察了花生四烯酸对TNF-α/放线菌素D(ActD)诱导内皮细胞凋亡的影响,现分析结果并探讨其作用机制。
1 材料与方法
1.1材料人脐静脉内皮细胞为本实验室冻存细胞株;内皮细胞培养液及生长因子、Annexin V-FITC试剂盒购自Life technologies。3-甲基腺嘌呤(3MA)、14,15-EET、微管相关蛋白轻链3-Ⅰ(LC3-Ⅰ)及微管相关蛋白轻链3-Ⅱ(LC3-Ⅱ)抗体、TNF-α、ActD购自Sigma公司;Caspse-8、Caspse-9活性检测试剂盒购自B&D公司。
1.2细胞培养采用含10% FBS的内皮细胞培养基培养人脐静脉内皮细胞,隔日换液,选用第4~5代细胞用于实验。将细胞随机分成空白组、模型组、EET组及EET+3MA组。空白组常规培养;模型组、EET组及EET+3MA组均加入TNF-α(10 ng/mL)和ActD(5 ng/mL)诱导凋亡;诱导凋亡的同时EET组加入14,15-EET(100 nmol/L); EET+3MA组加入14,15-EET (100 nmol/L)、3MA(2 mmol/L)。各组培养24 h时进行以下研究。
1.3细胞凋亡检测采用流式细胞仪。将上述各组细胞给予胰蛋白酶-EDTA消化,收集细胞至10 mL离心管中,低速离心后弃培养液。采用缓冲液漂洗细胞,调整细胞密度为1×106个/mL。取195 μL细胞悬液,加入PI和Annexin-v 5 μL,混匀后反应25 min。采用流式细胞仪检测细胞凋亡率。
1.4LC3-Ⅰ、 LC3-Ⅱ蛋白表达检测采用Western blotting法。将上述各组细胞采用冰PBS 洗涤,加入100 μL三去污细胞裂解液冰上裂解20 min;采用Bradford法测定样品蛋白含量,调整蛋白浓度;取相同总量的蛋白样品,加入加样缓冲液,高温凝固蛋白,加样至SDS聚丙稀酰胺中,持续电泳直至染料到底部;4 ℃、25 V电压下转膜过夜;封闭液浸泡封闭膜2 h,采用TBS-T液洗膜4次;分别加入LC3-Ⅰ、LC3-Ⅱ一抗(工作液浓度均为1∶3 000)孵育过夜;采用TBS-T液洗脱,加入辣根过氧化物酶标记的LC3-Ⅰ、LC3-Ⅱ二抗(工作液浓度均为1∶1 000),室温孵育1 h;采用TBS-T液洗膜3次,采用ECL化学发光法显影。采用Quantity One 图像分析软件对蛋白电泳条带的灰度值定量分析,计算目的蛋白相对表达量。目的蛋白相对表达量=目的蛋白灰度值/内参灰度值。
1.5Caspase-8、Caspase-9活性检测采用ELISA法。收集上述各组细胞,250 g离心10 min;加入裂解缓冲液,冰上孵育10 min;10 000 r/min离心1 min,收集上清;在酶标板中分别加入50 μL细胞裂解液、50 μL反应缓冲液、5 μL反应底物Caspase-8、Caspase-9;常温孵育1~2 h,采用酶标仪检测Caspase-8、Caspase-9活性,以光密度(OD)值表示。具体步骤参照Caspase-8、Caspase-9活性检测试剂盒说明书。
2 结果
2.1各组细胞凋亡率比较空白组、模型组、EET组及EET+3MA组细胞凋亡率分别为(0.22±0.11)%、(28.20±2.34)%、(8.23±1.14)%、(15.12±2.12)%,空白组、EET组及EET+3MA组细胞凋亡率均低于TNF-α组(P均<0.01),EET+3MA组高于EET组(P<0.05)。
2.2各组细胞LC3-Ⅰ、LC3-Ⅱ蛋白表达及LC3-Ⅱ/LC3-Ⅰ比较EET组及EET+3MA组LC3-Ⅱ/LC3-Ⅰ高于模型组、空白组(P<0.05或<0.01);EET+3MA组LC3-Ⅱ/LC3-Ⅰ低于EET组(P<0.01)。见表1。
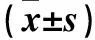
表1 各组细胞LC3-Ⅰ、LC3-Ⅱ蛋白表达
注:与空白组比较,#P<0.01;与TNF-α组比较,*P<0.05;与EET组比较,△P<0.01。
2.2各组细胞Caspase-8、Caspase-9活性比较模型组、EET组、EET+3MA组Caspase-8、Caspase-9活性均高于空白组,模型组高于EET+3MA组、EET组,EET+3MA组高于EET组,组间比较P<0.05或<0.01。见表2。

表2 各组细胞Caspase-8、Caspase-9活性比较
注:与空白组比较,#P<0.01;与TNF-α组比较,*P<0.05;与EET组比较,△P<0.01。
3 讨论
自噬是细胞利用溶酶体降解自身受损的细胞器和大分子物质的过程,是真核细胞特有的生命现象。研究表明,自噬现象在内皮细胞中广泛存在,抑制内皮细胞自噬能够促进氧化低密度脂蛋白诱导的内皮细胞损伤[4]。Chen等[5]报道,白藜芦醇通过上调自噬,减少TNF-α诱导的内皮细胞损伤;说明自噬是内皮细胞存活及保持活力的自我调节方式之一,适度的自噬可能会保护内皮细胞,抑制细胞凋亡。
花生四烯酸是内皮细胞富含的一种物质。近年研究发现,花生四烯酸可通过细胞色素P450(CYP)表氧化酶代谢产生四种EET,分别为5,6、8,9、11,12和14,15-EET。EET通过上调内皮细胞内源性一氧化氮合酶表达,促进NO释放,发挥舒张血管、保护内皮的生物学功能[8,9];采用CYP2J2转染或给予8,9-EET处理均可导致NF-κB、MMP-9表达下调,促进内皮细胞增殖和迁移,抑制主动脉平滑肌细胞增殖和迁移,发挥抗动脉粥样硬化的作用[10]。Dhanasekaran等[11]研究发现,心肌缺血处理20 min后应用11,12-EET、14,15-EET可显著缩小心肌梗死面积。上述研究提示,EET具有抑制内皮细胞凋亡及损伤,抑制平滑肌细胞增殖,保护心肌细胞等作用,对维持心血管系统稳定意义重大[2]。Yang等[3]在研究花生四烯酸抑制TNF-α诱导内皮细胞凋亡的过程中发现,花生四烯酸可能通过激活MAPK及PI3K/Akt等途径抗细胞凋亡;但抑制了MAPK及PI3K/Akt等途径并不能完全阻断花生四烯酸的这种抗凋亡作用,提示花生四烯酸抗凋亡仍可能存在其他机制。Samokhvalov等[12]证实,EET能上调自噬,抑制饥饿诱导的内皮凋亡,提示EET可能通过活化自噬途径发挥抗凋亡作用。
本研究采用联合TNF-α/ActD诱导的内皮细胞凋亡模型。TNF-α是内皮细胞凋亡研究中常用的诱导剂;ActD可抑制蛋白质或RNA合成,即抑制抗凋亡基因的表达;联合使用TNF-α/ActD能更好地诱导内皮细胞凋亡[13]。本研究结果显示,TNF-α/ActD诱导下内皮细胞出现了明显的凋亡,加入14,15-EET后内皮细胞凋亡减少,证实14,15-EET可抑制内皮细胞凋亡。研究发现,Beelinl/PI3K-Ⅲ复合体参与了自噬泡的形成和自噬的启动;而3MA可通过抑制PI3K-Ⅲ复合体形成阻滞自噬的发生[14]。证实3MA是一种常用的细胞自噬阻滞剂。本研究中EET+3MA组加入3MA处理后,14,15-EET的抗凋亡作用明显减弱,提示自噬可能在14,15-EET抗TNF-α诱导的内皮细胞凋亡中发挥作用。
Atg8是在酵母中发现的一种自噬发生必需的蛋白质分子[15]。LC3是哺乳动物中ATG8-PE共轭体的同源物,LC3-Ⅱ是细胞质中的LC3-Ⅰ经过剪切等处理后的功能片段,为自噬泡形成所必需,其含量与自噬水平相关。LC3-Ⅱ/LC3-Ⅰ是反映哺乳动物自噬水平公认的指标[16]。本研究结果显示,在TNF-α诱导下内皮细胞中LC3-Ⅰ向LC3-Ⅱ转化,提示TNF-α诱导下内皮细胞发生自噬,可能是内皮细胞对外界损伤的一种保护机制。加入14,15-EET后,LC3-Ⅱ表达升高,进一步说明14,15-EET促进自噬。3MA、14,15-EET同时处理的内皮细胞LC3-Ⅱ表达明显抑制,说明14,15-EET诱导的自噬参与了抗凋亡作用。提示内皮细胞通过激活自噬对抗外源性损伤。
尽管认为自噬与凋亡在形态学、发生机制等多个方面均存在不同,但越来越多的研究提示,自噬与凋亡二者之间相互联系及制约,其中关于Caspase家族与两者关联的研究最为广泛。有研究者利用RNA干扰技术下调Caspase-8表达以抑制细胞凋亡的发生,但是相比未干扰细胞,干扰细胞中出现了更多的自噬小体,说明RNA干扰技术抑制Caspase-8表达的同时激活了自噬途径[17]。Furuya等[18]发现自噬关键蛋白Beclin-1可通过提高Caspase-9活性而增加凋亡诱导剂引起的细胞凋亡,提示Caspase在自噬与凋亡交互作用中发挥重要作用。本研究TNF-α组细胞凋亡显著增加的同时,Caspase-8、Caspase-9活性均明显升高,提示二者在TNF-α诱导细胞凋亡中发挥作用。加入14,15-EET后二者活性明显下降,说明14,15-EET可能通过抑制Caspase-8、Caspase-9发挥抗凋亡作用。EET+3MA组同时给予14,15-EET和3MA可抑制细胞自噬,Caspase-8、Caspase-9活性再次明显升高,说明14,15-EET上调自噬的机制可能是抑制Caspase-8、Caspase-9活性。
综上所述,花生四烯酸可抑制TNF-α诱导的内皮细胞凋亡;通过抑制Caspase-8、Caspase-9活性诱导自噬可能是其作用机制。
参考文献:
[1] Winn RK, Harlan JM. The role of endothelial cell apoptosis in inflammatory and immune diseases[J]. J Thromb Haemost, 2005,3(8):1815-1824.
[2] Fleming I. The Pharma cology of the cytochrome P450 epoxygenase/soluble epoxide hydrolase axis in the vasculature and cardiovascular disease[J]. Pharmacol ReV, 2014,66(4):1106-1140.
[3] Yang SL, Lin L, Chen JX, et al. Cytochrome P450 epoxygenases protect endothelial cells from apoptosis induced by tumor necrosis factor-α via MAPK and PI3K/Akt signaling pathways[J]. Am J Physiol Heart Circ Physiol, 2007,293(1):142-151.
[4] Zhang YL, Cao YJ, Zhang X, et al. The autophagy lysosome pathway: a novel mechanism involved in the processing of oxidized LDL in human vascular endothelial cells[J]. Biochem Biophys Res Commu, 2010,394(2):377-382.
[5] Chen ML, Yi L, Jin X, et al. Resveratrol attenuates vascular endothelial inflammation by inducing autophagy through the cAMP signaling pathway[J]. Autophagy, 2014,10(10):1877.
[6] Zhang H, Park Y, Wu J, et al. Role of TNF-α alpha in vascular dysfunction[J]. Clin Sci, 2009,116(3):219-230.
[7] Nessler J, Nessler B, Kitliński M, et al. Concentration of BNP, endothelin, pro-inflammatory cytokines (TNF-alpha, IL-6) and exercise capacity in patients with heart failure treated with carvedilol[J]. Kardiol Pol, 2008,66(2):144-151.
[8] Jiang JG, Chen RJ, Xiao B, et al. Regulation of endothelial nitric-oxide synthase activity through phosphorylationin response to epoxyeicosatrienoic acids[J]. Prostaglandins Other Lipid Mediat, 2007,82(1-4):162-174.
[9] Hercule HC, Schunck WH, Gross V, et al. Interaction between P450 eicosanoids and nitric oxide in the control of arterial tone in mice[J]. Arterioscler Thromb Vasc Biol, 2009,29(1):54-60.
[10] Schuck RN, Theken KN, Edin ML, et al. Cytochrome P450-derived eicosanoids and vascular dysfunction in coronary artery disease patients[J]. Atherosclerosis, 2013,227(2):442-448.
[11] Dhanasekaran A, Gruenloh SK, Buonaccorsi JN, et al. Multiple antiapoptotic targets of the PI3K/Akt survival pathway are activated by epoxyeicosatrienoic acids to protect cardiomyocytes from hypoxia/anoxia[J]. Am J Physiol Heart Circ Physiol, 2008,294(2):724-735.
[12] Samokhvalov V, Alsaleh N, El-Sikhry HE, et al. Epoxyeicosatrienoic acids protect cardiac cells during starvation by modulating an autophagic response[J]. Cell Death Dis, 2013(4):e885.
[13] 林立,王红.TNF-α/放线菌素D诱导原代内皮细胞凋亡模型的建立[J].中国分子心脏病学杂志,2006,6(6):332-337.
[14] Klionsky DJ, Abdalla FC, Abeliovich H, et al. Guidelines for the use and interpretation of assays for monitoring autophagy[J]. Autophagy, 2012,12(1):445-544.
[15] Nakatogawa H, Lehimura Y, Ohsumi Y. Atg8, a ubiquitin-like protein required for autophagosome formation, mediates membrane tethering and hemifusion[J]. Cell, 2007,130(1):165-178.
[16] Johansen T, Lamark T. Selective autophagy mediated by autophagic adapter proteins[J]. Autophagy, 2011,7(3):279-296.
[17] Lu C. Autophagic degradation of active caspase-8: a cross talk mechanism between autophagy and apoptosis[J]. Autophagy, 2010,6(7):891-900.
[18] Furuya D, Tsuji N, Yagihashi A, et al. Beclin-1 augmented cis-diamminedichloroplatinum induced apoptosis via enhancing caspase-9 activity[J]. Exp Cell Res, 2005,307(1):26-40.
猜你喜欢
儿童时代·快乐苗苗(2022年10期)2022-12-09
中国眼镜科技杂志(2019年9期)2019-11-11
水产科学(2018年2期)2018-03-27
安徽医科大学学报(2016年12期)2017-01-15
儿童时代·快乐苗苗(2016年4期)2016-11-07
学苑创造·A版(2016年7期)2016-07-06
中国粮油学报(2016年5期)2016-01-23
中国粮油学报(2016年5期)2016-01-23
中国病理生理杂志(2015年8期)2015-12-21
云南中医学院学报(2014年5期)2014-07-31
