
两个春小麦品系抗叶锈性遗传分析
2018-05-11 09:36袁军海沈凤英吴伟刚张爱香
麦类作物学报 2018年4期
袁军海,沈凤英,吴伟刚,张爱香
(河北北方学院植物保护系,河北宣化 075131)
叶锈病是小麦的主要病害之一,一般会导致小麦减产5%~10%,流行年份可导致减产30%以上。此类病害主要通过培育抗病品种和喷洒化学药剂防治。但由于小麦生产经济效益较低,加之化学防治易造成环境和农产品污染等,所以农民更倾向于选用抗病品种。遗传分析是抗病品种选育的基础,但到目前为止,我国对小麦抗叶锈病遗传分析研究较少[1-2],或未经F3或测交F2验证,结果可信度不高[3-4],背景尚不明确。8821-1-1由甘肃省定西市旱作农业科研推广中心选育,系谱为“RFMⅢ-87-A/宁春10号”[5];九三98-61297由黑龙江省农垦总局九三科学研究所选育,系谱为“(西引1号/41123)F1/多父本”[6]。两品系均属春性,综合农艺性状较好,2006年经中国农业科学院植物保护研究所田间抗叶锈病鉴定,前者严重度为10%,且表现中度抗病至高度抗病,后者表现免疫,均有较大应用价值。本试验对二者的抗叶锈性进行了遗传分析。
1 材料与方法
1.1 试验材料
春小麦品系8821-1-1和九三98-61297由育种单位提供,小麦抗叶锈病基因 Lr13的载体品系Manitou和Tc*6/Manitou,及感病品种Thatcher,均由国际玉米小麦改良中心(CIMMYT, Centro Internacional de Mejoramiento de Maizy Trigo)提供。小麦叶锈病菌致病类型PHT/RN、PHT/RP和THT/TP,由中国农业科学院植物保护研究所提供,均采自我国各小麦产区,按Long等[7]和Singh[8]提出的密码法则命名。
1.2 试验方法
配制以下杂交组合:Thatcher×8821-1-1、Manitou×8821-1-1、Thatcher×九三98-61297、Manitou×九三98-61297、8821-1-1×九三98-61297。获得F1后,每个杂交组合随机选取5粒种子播种,花前套袋自交,所获得的5株的种子混合作为F2群体,再从中随机选取180粒种子播种并套袋自交,每粒种子的后代作为1个F3株系。
苗期抗病性测定在塑料盒内进行,塑料盒规格为35 cm×20 cm×10 cm,预先放入营养土。供试品种(系)种子5粒、F1种子10粒、F2种子400~600粒,均穴播,每穴5~7粒,穴深1 cm,穴行、穴距均5 cm;每个F3株系65~70粒种子,条播,深1 cm,长9 cm。待麦苗第一叶片充分展开后,先对叶片用清水去蜡,然后喷洒各致病类型的夏孢子悬浮液(供试品种或品系用PHT/RN、PHT/RP和THT/RP,杂交后代用PHT/RP)、放入保湿桶内,在室温条件下黑暗保湿16 h,然后转入RXZ-280B型人工气候箱(宁波江南仪器厂生产)中培养,每天光照14 h,光照强度12 000 Lx。设置18 ℃ 和28 ℃两种温度条件。28 ℃ 条件下8~10 d后、18 ℃ 12~14 d后,根据侵染型分级标准[9]调查(28 ℃下THT/TP侵染型未调查)。将0~X归为抗病类型,3~4归为感病类型,进行遗传分析和杂交验证。
成株期抗病性测定在河北北方学院南校区农场进行。小区规格11 m×2.2 m。行距0.33 m,每小区34行;两端为保护行,每隔10行设1个诱发行,即保护行和诱发行各2行;剩余的30行均横向分为3部分:左、右各1 m(称为“半行”)播种待鉴定材料,中间0.2 m与各行垂直方向纵向播种1行诱发行;保护行和诱发行品种均为Thatcher。供试品种(系)约50粒种子,均条播1个“半行”;F110粒种子、F2400~700粒种子,均点播,株距10 cm;每个F3株系65~70粒种子,条播1个“半行”。4月中旬约小麦起身期,对诱发行接种。每3 m设1个接种点。成株期抗病性测定所用菌种为PHT/RN、PHT/RP和THT/TP三种致病类型的等比例混合菌种。选晴天无风的傍晚,先对麦苗基部浇水至土壤含水量饱和,然后用清水对上部叶片去蜡、喷洒孢子悬浮液,盖膜保湿12~14 h后揭膜。约7月中旬,待感病对照品种Thatcher旗叶叶锈病严重度达60%~80%时,以旗叶为准,调查待鉴定材料各植株的抗感反应和严重度。抗感反应根据Roelfs[9]确定的侵染型标准调查后判断,基本等级为免疫(immunity,I)、抗病(resistant,R)、中度抗病(moderately resistant,MR)、中度感病(moderately susceptible,MS)和感病(susceptible,S);严重度根据Peterson等[10]提出的“改良Cobb标准”判断。由于工作量较大,F3仅判断整个株系的抗病、感病情况,未做单株调查。
2 结果与分析
2.1 供试材料对叶锈病的抗性
除在18 ℃对叶锈病菌致病类型PHT/RP和THT/TP表现高度感病外,8821-1-1在苗期均表现中度抗病,在成株期表现为严重度10%且中度抗病至抗病;除在18 ℃ 对致病类型PHT/RN和28 ℃对致病类型PHT/RP表现中度抗病外,九三98-61297在苗期均表现感病,在成株期表现免疫(表1)。 Lr13的载体品系Manitou在苗期、18 ℃ 对3种致病类型均高度感病,在28 ℃ 对致病类型PHT/RN和PHT/RP均表现中度抗病,在成株期表现为严重度40%且抗病;另一载体品系Tc*6/Manitou苗期仅在28 ℃ 对致病类型PHT/RP表现中度抗病,其他情况均感病或高度感病,成株期表现免疫(表1)。Thatcher在苗期和成株期均高度感病(表1)。
2.2 苗期抗叶锈性遗传分析
在18 ℃,所有杂交组合的所有世代均表现高度感病。在28 ℃,Thatcher×8821-1-1杂交组合,6株F1植株均表现为中度抗病,378株F2植株中,271株为抗病类型、107株为感病类型,能够符合3抗病∶1感病的期望比例,故推定8821-1-1含有1对显性抗叶锈病基因;135个F3株系中,29个株系全部抗病、73个株系抗感分离、33个株系全部感病,能够符合1全部抗病∶2抗感分离∶1全部感病的期望比例,可以验证8821-1-1含有1对显性抗叶锈病基因(表2)。相同条件下,对(Manitou×8821-1-1)F2代552个单株进行测定,全部为抗病类型,说明8821-1-1含有 Lr13(表2)。综合判断,在28 ℃下,Thatcher×8821-1-1杂交组合中对致病类型PHT/RP具有抗性的一对显性基因是 Lr13。同理可判断,九三98-61297也可能含有 Lr13。相同条件下,8821-1-1×九三98-61297杂交组合的F2共测定423株,全部为抗病类型,也可验证两品系所含的有效基因相同(表2)。
表1供试材料苗期和成株期的抗叶锈性
Table1LeafrustresistanceoftestedmaterialstoPucciniatriticinaatseedlingandadult-plantstages

供试材料Testedmaterial苗期抗病性SeedlingresistancePHT/RN18℃28℃PHT/RP18℃28℃THT/TP18℃成株期抗病性Adult⁃plantresistance严重度Severity/%抗感反应Resistantresponse8821⁃1⁃11+3;12+3+;1333+10MRR九三98⁃612972+33+3;3+0IManitou33+;1+33+;12+3+40RTc∗6/Manitou3+33+X+3+0IThatcher3+3+3+3+3+≥80S
MRR: 中度抗病至抗病; I:免疫;R:抗病;S:感病;“;1”指同一叶片随机出现“;”和“1”两种侵染型,但以第一种为主,其余情况类推;“X+”指同一叶片随机出现所有侵染型但以感病类型为主。
MRR:Moderate to highly resistance; I:Immunity; R:Resistant;S:Susceptible“;1”indicates random distribution of two infection types;“;” and “1” on single leaf,with the most prevalent infection type listed first,and similarly as to the others;“X+” indicates random distribution of all infection types on single leaf,and the susceptible infection type predominate.
表2供试材料苗期抗叶锈性的遗传分析
Table2Geneticanalysisofleafrustresistanceatseedlingstageintestedmaterials

杂交组合Crosscombination世代Generation侵染型分布Distributionofinfectiontype0;;112X33+4总株数No.oftotalplants植株或株系数No.ofplantsorlines抗病Resistant分离Segregate感病Susceptible理论比例Expectedratioχ2值χ2valueP值PvalueThatcher×8821⁃1⁃1F1666-0F2111532424364378271-1073∶12.030.10~0.25F312144620311991177229--1∶2∶11.130.50~0.75259639991320543476984109-73-64711781825--33Manitou×8821⁃1⁃1F21658223453552552-01∶0Thatcher×九三98⁃61297F1777-0F237172474287457328-1293∶12.370.10~0.25F31287851752205335--1∶2∶11.760.25~0.505814328252655412574989-84-67218612533--47Manitou×九三98⁃61297F2359234459585585-01∶08821⁃1⁃1×九三98⁃61297F21610397423423-01∶0
F3代浸染型分布情况分3行著录,第1行、第2行、第3行分别为全部抗病、抗感分离和全部感病株系各单株。
The distribution of F3infection types were listed in three lines, including plants of homozygous resistant families,segregating families and homozygous susceptible families,respectively.
2.3 成株期抗叶锈性遗传分析
Thatcher×8821-1-1后代中,F1测定6株,5株的严重度低于30%,1株的严重度为31%~60%,但均表现中度抗病至高度抗病(表3)。F2测定427株,303株表现中度抗病至高度抗病,严重度从0至80%以上不等,124株表现中度感病至高度感病,3株的严重度低于30%,121株的严重度高于60%(表3)。若不考虑严重度,303株抗病,124株感病,符合3抗病∶1感病的期望比例;若考虑严重度,由于表现中度感病至高度感病且严重度低于30%,可视为慢锈,仍有较大应用价值[11],可将表现中度感病至高度感病且严重度低于30%的3个植株归为抗病,则总计306株抗病、121株感病,也符合3抗病∶1感病的期望比例。结合F1,可推定8821-1-1含有1对显性抗叶锈病基因。F3测定152个株系,若按上述抗感标准判断,则有33个株系全部抗病,71个株系抗感分离,48个株系全部感病,能够符合1全部抗病∶2抗感分离∶1全部感病的期望比例,可以验证8821-1-1含有1对显性抗叶锈病基因(表3)。相同条件下,Manitou×8821-1-1杂交F2代共测定687株,若按上述抗感标准判断,则全部抗病,说明8821-1-1含有 Lr13基因。综合判断,8821-1-1 在成株期的抗叶锈性由 Lr13基因控制。与苗期相同,同理可判断:九三98-61297在成株期的抗叶锈性也由 Lr13控制,8821-1-1和九三98-61297所含在成株期有效的抗叶锈病基因相同。Thatcher×九三98-61297杂交组合的F2,既符合3抗∶1感的期望比例,也符合13抗∶3感的期望比例,但F3仅符合1全抗∶2分离∶1全感的期望比例,故否定后一种可能性(表3)。
表3供试材料成株期抗叶锈性遗传分析
Table3Geneticanalysisofleafrustresistanceatadult-plantstageintestedmaterials
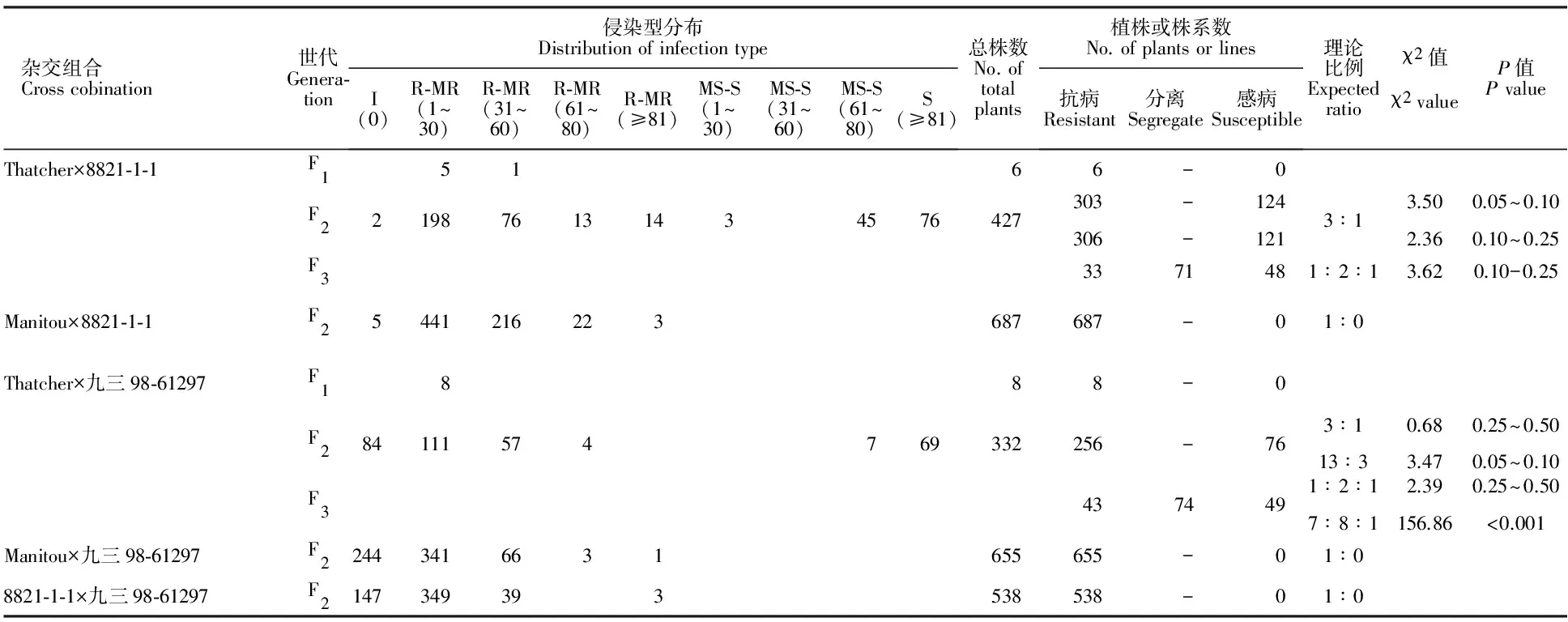
杂交组合Crosscobination世代Genera⁃tion侵染型分布DistributionofinfectiontypeI(0)R⁃MR(1~30)R⁃MR(31~60)R⁃MR(61~80)R⁃MR(≥81)MS⁃S(1~30)MS⁃S(31~60)MS⁃S(61~80)S(≥81)总株数No.oftotalplants植株或株系数No.ofplantsorlines抗病Resistant分离Segregate感病Susceptible理论比例Expectedratioχ2值χ2valueP值PvalueThatcher×8821⁃1⁃1F15166-0F2219876131434576427303306--1241213∶13.502.360.05~0.100.10~0.25F33371481∶2∶13.620.10-0.25Manitou×8821⁃1⁃1F25441216223687687-01∶0Thatcher×九三98⁃61297F1888-0F284111574769332256-763∶113∶30.683.470.25~0.500.05~0.10F34374491∶2∶17∶8∶12.39156.860.25~0.50<0.001Manitou×九三98⁃61297F22443416631655655-01∶08821⁃1⁃1×九三98⁃61297F2147349393538538-01∶0
I: 免疫; R-MR: 抗病至中度抗病; MS-S:中度感病至感病; S:感病;各侵染型后的括号中所括数字为严重度,单位为%。
I: Immunity; R-MR: Resistant to moderately resistant; MS-S: Moderately susceptible to susceptible; S: Susceptible;The number in the parenthes following each infection type is severity,and the unit is %.
3 讨 论
袁军海等[6]通过基因推导认为,8821-1-1含有 Lr13和未知基因,而九三98-61297含有未知基因,未推导出 Lr13。本试验通过杂交验证证实,两品系均含有 Lr13。反观基因推导结果,可能与 Lr13不易在苗期表达,且其表达受遗传背景影响较大有关,如苗期在28 ℃,载体品系Manitou对PHT/RP等5个致病类型表现中度抗病,而Tc*6/Manitou则仅对PHT/RP中度抗病,其余均高度感病。若以Tc*6/Manitou为标准,可推断九三98-61297含有 Lr13,且二者在成株期均表现免疫,但这样的推导结果可信度并不高。基因推导结果不能完全通过等位性杂交来验证的现象,其他文献也有类似报道[12-14]。所以,本试验的遗传分析和杂交验证,也可视为与基因推导结果间的相互验证。总之,将遗传分析与杂交验证和基因推导等方法结合起来,才能提高结果的可信度。
在我国小麦品种所含主要抗叶锈病基因中, Lr13的出现频率约为5.4%,目前仍比较有效[15]。主要表现为:(1) Lr13可提高其他抗叶锈病基因的抗病性。如Kolmer[16]发现, Lr1、Lr3、Lr11、Lr24、Lr26等基因,单独应用时在成株期已基本丧失抗病性(≥70MS),但与 Lr13组合起来,抗病性明显提高(10~40MR),且组合的抗病性也高于 Lr13单独存在时的抗病性(20~60MR)。(2) Lr13主要在成株期表达,具有慢锈性,进而具有持久抗叶锈性潜质[16-18]。但值得注意的是, Lr13对致病类型也有专化性,当遇到毒性致病类型时, Lr13的慢锈性会失效。如对来自印度的小种77-A, Lr13在整个生育期均表现感病[19];用来自墨西哥的致病类型TCB/TD等测定,许多含有 Lr13的品种(系)在成株期均高度感病[20-23];类似现象在用来自澳大利亚的致病类型测定时也有发现[24]。我国目前尚无明确的研究报道。(3) Lr13具有高温抗病性[25]。如在袁军海等[6]的试验中, Lr13对随机选取的PHT等4个致病类型均表现为高温抗病性。PHT一直是我国小麦叶锈病菌的优势致病类型[26-27]。在我国广大冬麦区,叶锈病逐渐流行时也是温度逐渐增高时,恰恰适宜 Lr13的表达。这可能是 Lr13在我国目前尚比较有效的主要原因[15,28]。故进行抗病育种时,应尽量将 Lr13与其他抗叶锈病基因组合到同一品种中;进行品种布局时,建议将含有 Lr13的抗病品种安排在小麦生长后期温度较高的冬麦区,更易发挥 Lr13的抗病作用。
4 结 论
8821-1-1和九三98-61297均含有 Lr13,在苗期28 ℃ 控制对致病类型PHT/RP的抗性,在成株期控制对PHT/RP等3种致病类型等比混合菌种的抗性,且均呈显性遗传。
致谢:中国农业科学院植物保护研究所的刘太国研究员和冯晶副研究员参与部分工作,谨此致谢!
参考文献:
[1] 赤国彤,王焕如,朱之堉.7个小麦品种抗叶锈病遗传研究初报[J].河北农业大学学报,1991,14(4):76.
CHI G T,WANG H R,ZHU Z Y.Genetic analysis of leaf rust resistance in seven varieties of wheat [J].JournalofHebeiAgriculturalUniversity,1991,14(4):76.
[2] 袁军海,陈万权.春小麦品种青春221抗叶锈性遗传分析[J].植物保护学报,2013,40(1):20.
YUAN J H,CHEN W Q.Genetic analysis of leaf rust resistance in spring wheat Qingchun 221 [J].ActaPhytophylacicaSinica,2013,40(1):20.
[3] 张 庆,刘太国,张 敏,等.小麦抗源Sw92抗叶锈病基因遗传及其分子标记[J].植物保护学报,2007,34(5):515.
ZHANG Q,LIU T G,ZHANG M,etal.Genetics and SSR markers of gene for leaf rust resistance in winter wheat line SW92 [J].ActaPhytophylacicaSinica,2007,34(5):515.
[4] 张培培,周 悦,董海焦,等. 周麦11、西农1163-4抗叶锈病基因与周8425B中 LrZH84 的关系[J].江苏农业科学,2015,43(10):33.
ZHANG P P,ZHOU Y,DONG H J,etal.Relationship between leaf rust resistance genes in Zhoumai 11 and Xinong 1163-4 and LrZH84 in Zhou 8425B [J].JournalofJiangsuAgriculturalSciences,2015,43(10):33.
[5] 牟丽明.旱地春小麦新品系8821-1-1选育报告[J].甘肃农业科技,2007(12):9.
MOU L M.Breeding report on the novel spring wheat line 8821-1-1 in dryland[J].GansuAgriculturalScienceandTechnology,2007(12):9.
建筑物的围护结构一般由屋顶、墙、地基隔热材料、密封材料、门和窗以及遮阳设施构成。一般情况下,增加建筑围护结构的费用仅占总投资额的3%~6%,节能却高达20%~40%。构建建筑物的保温隔热系统是对建筑物采取良好的保温隔热的主要措施。外墙外保温是严寒地区保温效果最好、应用最广泛的一种建筑保温形式。保温材料选择主要考虑导热率与燃烧特性,欧盟新研制的只允许单方向传热的热二极管墙体可以产生良好的隔热效果,也可以运用具有性价比高的热工性能玻璃。外墙保温厚度由所在气候区一级基层墙体材料所控制,外墙保温隔热层厚度一般为23~40cm,屋顶保温采用隔热方式,也可采用反射降温等方式。
[6] 袁军海,刘太国,陈万权.中国47个小麦新品种(系)苗期抗叶锈病基因推导[J].中国农业科学,2007,40(9):1925.
YUAN J H,LIU T G,CHEN W Q.Postulation of leaf rust resistance genes in 47 new wheat cultivars(lines) at seedling stage [J].ScientiaAgriculturaSinica,2007,40(9):1925.
[7] LONG D L,KOLMER J A.A north American system of nomenclature forPucciniareconditaf.sp.tritici[J].Phytopathology,1989,79(5):525.
[8] SINGH R P.Pathogenicity variations ofPucciniareconditaf.sp.triticiandP.graminisf.sp.triticiin wheat-growing areas of Mexico during 1988 and 1989 [J].PlantDisease,1991,75:790.
[9] ROELFS A P.Race specificity and methods of study [M]//BUSHNELL W R,ROELFS A P.The cereal rust Ⅰ.origins,specificity,structure,and physiology.New York:Academic Press,1985:134.
[10] PETERSON R F,CAMPBELL A B,HANNAH A E.A diagrammatic scale for estimating rust intensity on leaves and stems of cereals [J].CanadianJournalofResearch(SectionC),1948,26:496.
[11] 陈万权,秦庆明,陈扬林,等.小麦成株期慢叶锈性初步筛选[J].西南农业学报,1998,11(1):54.
[12] MODAWI R S,BROWDER L E,HEYNE E G.Use of infection-type data to identify genes for low reaction toPucciniareconditain several winter wheat cultivars [J].CropScience,1985,25:9.
[13] MODAWI R S,BROWDER L E,HEYNE E G.Genes for low reaction toPucciniareconditain Newton hard red winter wheat [J].CropScience,1985,25:13.
[14] MCVEY D V.Verification of infection-type data for identification of genes for resistance to leaf rust in some hard red spring wheat [J].CropScience,1989,29:304.
[15] 袁军海,陈万权.中国小麦主要抗叶锈病基因的有效性评价[J].麦类作物学报,2011,35(7):794.
YUAN J H,CHEN W Q.Estimate on the effectiveness of main resistant genes for leaf rust in Chinese wheat [J].JournalofTriticeaeCrops,2011,35(7):794.
[16] KOLMER J A.Enhanced leaf rust resistance in wheat conditioned by resistance gene pairs with Lr13[J].Euphytica,1992,61:123.
[17] SAMBORSKI D J,DYCK P L.Enhancement of resistance toPucciniareconditaby interaction of resistance genes in wheat [J].CanadianJournalofPlantPathology,1982,4:152.
[18] 陈万权,秦庆明.国际上已知小麦抗叶锈病基因在中国的可利用性研究[J].中国农业科学,2002,35(7):794.
CHEN W Q,QIN Q M.Studies on utilization of worldwide known genes for leaf rust resistance of wheat in China [J].ScientiaAgriculturaSinica,2002,35(7):794.
[19] SAINI R G,GUPTA A K,ANAND D.Expression of some leaf rust resistance genes at different growth stages in wheat against race 77-A [J].CurrentScience,1986,55:802.
[20] SINGH R P,GUPTA A K.Genes for leaf rust resistance in Indian and Pakistani wheats tested with Mexican pathotypes ofPucciniareconditaf.sp.tritici[J].Euphytica,1991,57:27.
[21] SINGH R P,RAJARAM S.Resistance toPucciniareconditaf.sp.triticiin 50 Mexico bread wheat cultivars [J].CropScience,1991,31:1472.
[22] SINGH R P.Association between gene Lr34 for leaf rust resistance and leaf tip necrosis in wheat [J].CropScience,1992,32:874.
[23] SINGH R P.Resistance to leaf rust in 26 Mexico wheat cultivars [J].CropScience,1993,33:633.
[24] SINGH D,PARK R F,MCINTOSH R A.Postulation of leaf (brown) rust resistance genes in 70 wheat cultivars grown in the United Kingdom [J].Euphytica,2001,120:205.
[25] PRETORIUS Z A,WILCOXSON R D,LONG D L,etal.Detecting wheat leaf rust resistance gene Lr13 in seedling [J].PlantDisease,1984,68:585.
[26] 葛润静,刘太国,高 利,等.2011-2012年度中国六省小麦叶锈菌群体毒性分析[J].植物病理学报,2015,45(2):175.
GE R J,LIU T G,GAO L,etal.Virulence ofPucciniatriticinafrom 6 provinces in China in 2011-2012 [J].ActaPhytopathologicaSinica,2015,45(2):175.
[27] 原宗英,武英鹏,刘敏捷.山西省小麦叶锈菌致病类型及毒性监测[J].山西农业科学,2017,45(3):448.
YUAN Z Y,WU Y P,LIU M J.Pathogenic types and virulence of wheat leaf rust (Pucciniatriticina) in Shanxi Province [J].JournalofShanxiAgriculturalSciences,2017,45(3):448.
[28] 闫晓翠,李在峰,杨华丽,等.30个重要小麦生产品种抗叶锈性基因分析[J].中国农业科学,2017,50(2):272.
YAN X C,LI Z F,YANG H L,etal.Analysis of wheat leaf rust resistance genes in 30 important wheat cultivars [J].ScientiaAgriculturaSinica,2017,50(2):272.
猜你喜欢
西南农业学报(2022年4期)2022-05-28
江西农业学报(2021年8期)2021-09-08
湖北农业科学(2021年10期)2021-06-25
中国蔬菜(2021年3期)2021-03-29
世界农药(2020年4期)2020-12-22
农业科技与信息(2020年20期)2020-12-18
上海师范大学学报·自然科学版(2020年5期)2020-12-18
贵州农业科学(2020年1期)2020-03-25
植物研究(2020年6期)2020-03-05
上海农业科技(2019年3期)2019-06-25
