
淇河鲫不同家系生长性能的比较
2018-05-17 07:19施文瑞宋东蓥吴利敏李学军陈军平张世杰孙亚威许莎诺
淡水渔业 2018年3期
王 磊,施文瑞,宋东蓥,马 晓,吴利敏,李学军,陈军平,张世杰,孙亚威,许莎诺
(河南师范大学水产学院,河南新乡 453007)
淇河鲫(Carassiusauratusvar.Qihe),属鲤形目(Cypriniformes)鲤科(Cyprinidae)鲫属(Carassius)[1-2],是天然雌核发育三倍体鲫[3-6],属于银鲫的一种[7],具有天然雌核发育和有性生殖两种生殖方式[8]。淇河鲫因原产于河南省淇河,故称“淇河鲫”,其肉质鲜美,自古就有“以其食鱼,唯淇之鲫”的记载,在古代曾被列为贡品[9],属于河南省特有的名贵地方鱼类。有研究表明淇河鲫的必需氨基酸占总氨基酸的百分比和必需氨基酸指数等都高于青鱼(Mylopharyngodonpiceus)、草鱼(Ctenopharynodonidellus)、鲢(Hypophthalmichthysmolitrix)、鳙(Aristichthysnobilis)和鲤(Cyprinuscarpio)等鱼类[10],与彭泽鲫(C.auratusvar.Pengze)相比,其含肉率和蛋白含量都较高[11],与普通野生鲫(C.auratus)相比,淇河鲫在体高和体厚等方面均显著高于野生鲫[3]。
虽然淇河鲫具有较高的营养价值和经济价值,然而随着其生存环境的恶化和野生资源的衰退[2],淇河鲫养殖产业的发展也受到了重大威胁,因此有必要开展对淇河鲫的选育工作。虽然我国的水产研究已经进入了基因组时代[12],并先后完成了半滑舌鳎(Cynoglossussemilaevis)[13]、大黄鱼(Pseudosciaenacrocea)[14]、鲤[15]和牙鲆(Paralichthysolivaceus)[16]等的全基因组测序工作,且将其成果应用于遗传育种研究,但家系选育仍是目前鱼类新品种培育中应用最多的方法之一[17],马爱军等[18]利用鳗弧菌(Vibrioanguillarum)对大菱鲆(Scophthalmusmaximus)30个家系进行感染实验,从中筛选出5个抗病力较强的家系;王磊等[19]对38个牙鲆家系的4个选育性状进行了分析,不仅揭示了4个性状之间的相关性,而且筛选到了4个抗病家系和7个高养殖存活率的家系;王兰梅等[20]通过对福瑞鲤家系不同养殖阶段的生长差异进行分析,发现不同月龄的体质量相关性均达到极显著水平,因此在选育过程中进行早期选择是有必要的。本研究利用4个鲤群体和1个淇河鲫群体建立若干淇河鲫家系,通过测量其生长性状和统计养殖成活率,以期筛选出生长速度快、体型好和养殖成活率高的家系,为淇河鲫优良品系的培育提供理论与实践依据。
1 材料与方法
1.1 亲鱼来源
实验所用的雌性亲本为河南师范大学保存的淇河鲫群体,雄性亲本分别为超级鲤群体、河南省黄河鲤鱼良种场黄河鲤群体(位置位于武陟县詹店镇,故简称“詹店黄河鲤”)、河南省水产良种繁育场黄河鲤群体(简称“良种场黄河鲤”)、楚楼水库黄河鲤群体和河南师范大学保存的淇河鲫群体。亲鱼的养殖和繁殖等均在河南师范大学水产养殖基地完成。
1.2 家系建立
用于建立家系的淇河鲫群体均为2龄鱼,黄河鲤和超级鲤雄鱼均为1龄鱼,从亲本中选取健康个体采用干法受精的方法在23 ℃的水温下共建立了19个家系(见表1),其中1~20号家系受精日期为2015年5月9号,22~33号家系受精日期为2015年5月14号。19个家系中互为母本半同胞家系有:10号和14号,11号和15号,32号和33号;互为父本半同胞家系有:1号和2号,10号、11号和12号,13号、14号和15号,20号和22号,26号和27号。

表1 家系亲本组合Tab.1 The parent composition of families
注:QH代表淇河鲫群体;SC代表超级鲤群体;LZC代表良种场黄河鲤;CL代表楚楼水库黄河鲤群体;ZD代表詹店黄河鲤。表2、表5同。
1.3 家系培育及生长测量
先后采取环境条件标准化和数量标准化的方式对家系进行培养以减小养殖环境差异对实验结果的影响。仔鱼孵化后转入大小一致的塑料桶进行培育,培育6个月后从每个家系中随机选取50尾注射PIT(Passive Integrated Transponder)标记。
家系养殖22个月后,对标记的实验鱼分别扫码并测量其体长、体高和体质量,其中体高与体长精确到毫米,体质量精确到克。
1.4 数据分析
存活率计算公式为:存活率=100%×终末鱼数/初始鱼数[21]
利用SPSS 16.0软件分别对家系的各个性状进行单因素方差分析、LSD多重检验分析和Pearson相关性分析。运用Excel计算不同家系各个性状的变异系数。
2 结果与分析
2.1 家系存活率比较
从6~22月龄,19个家系的存活率在36%~86%之间(表2),平均存活率为64%,存活率高于80%的家系有5个,分别为7号、10号、13号、14号和15号家系,虽然这5个家系的父本均不是淇河鲫,但是父本不是淇河鲫的家系中也有存活率比较低的,如2号、11号和26号家系(存活率均为42%),因此存活率的高低与父本是淇河鲫还是鲤无必然联系。另外,13号、14号和15号家系为互为父本半同胞家系,显示由这条鱼作为父本的后代可能具有较高的存活率,因此存活率的高低可能与具体的父母本有关。

表2 19个淇河鲫家系的养殖存活率Tab.2 Breed survival rate of 19 C.sauratus var.Qihe families
2.2 家系生长性状的比较
19个家系22月龄体长、体高和体质量值见表2,体长平均值为(14.35±1.23) cm,范围13.57~16.07 cm;体高平均值为(5.13±0.52) cm,范围3.1~7.9 cm;体质量平均值为(91.68±23.62) g,范围75.03~130.81 g。
以各性状表型值最接近均值的12号家系为对照组进行单因素方差分析,2号、11号、18号、22号和32号家系体长与对照组相比差异极显著,7号、26号和33号家系与对照组相比差异显著;2号、11号、22号和33号家系体高与对照组相比差异极显著,7号、10号、26号、29号和32号家系与对照组相比差异显著;2号、7号、11号、18号、22号、32号和33号家系体质量与对照组相比差异极显著,13号、24号、26号和29号与对照组相比差异显著(表3)。

表3 不同家系各生长性状的表型测量值Tab.3 Growth traits in different families

续表3
注:以各性状表型测量值均接近均值的12号家系为对照组,**表示差异极显著,*表示差异显著
对体长、体高与体质量进行相关性分析(表4),结果显示各个性状间的相关性均为极显著正相关,因此将体质量作为主要选育指标,结合各家系的体长、体高与体质量的表型值及LSD多重检验结果,将2号和11号家系定义为快速生长家系,18号、24号、26号与29号家系定义为生长较快的家系。

表4 淇河鲫各生长性状相关性比较Tab.4 Correlation coefficient between different growth traits
注:**表示在0.01水平(双侧)上显著相关
2.3 不同家系体型的比较
在鲫的选育过程中,一般认为高背型经济价值更高,因此对各个淇河鲫家系的体高/体长进行了分析,19个家系体高与体长的比值见表5,各个家系体高与体长的比值从0.347到0.367不等,超过0.365的家系有2号和29号家系。
2.4 不同家系变异系数分析
不同家系各性状的变异系数见表6,19个家系体长变异系数平均值为8.6%,范围2.8%~11.7%;体高变异系数平均值为10.2%,范围4.9%~14.1%;体质量变异系数平均值为25.8%,范围6.4%~31.7%。综合3个性状的变异系数,24号、32号与33号家系的变异系数均较小,说明其遗传稳定性较高;11号、13号、14号、15号和22号家系各性状变异系数较高,可以认为其遗传多样性较大,具有较大的选择潜力[12]。
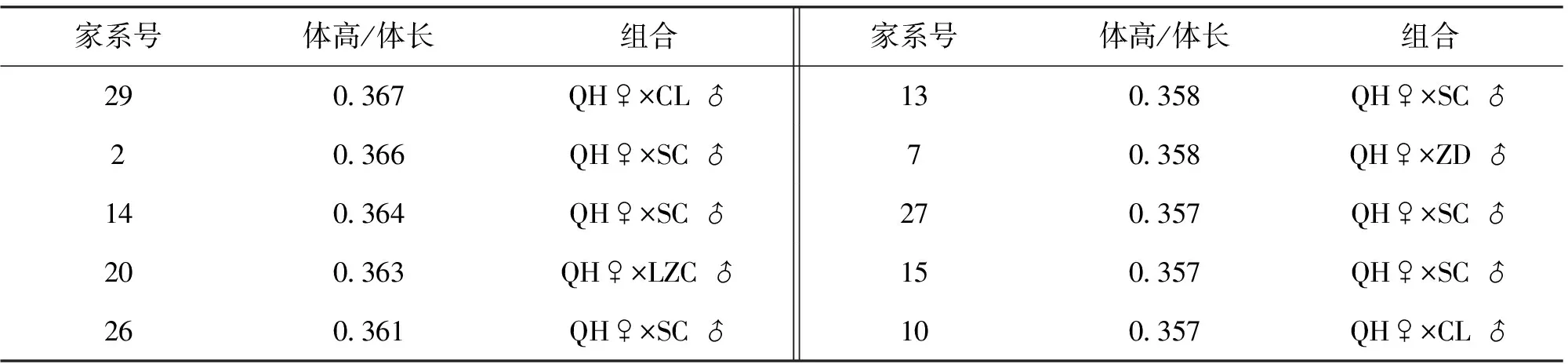
表5 淇河鲫家系体型比较Tab.5 Comparison of bodily form between C.sauratus var.Qihe families
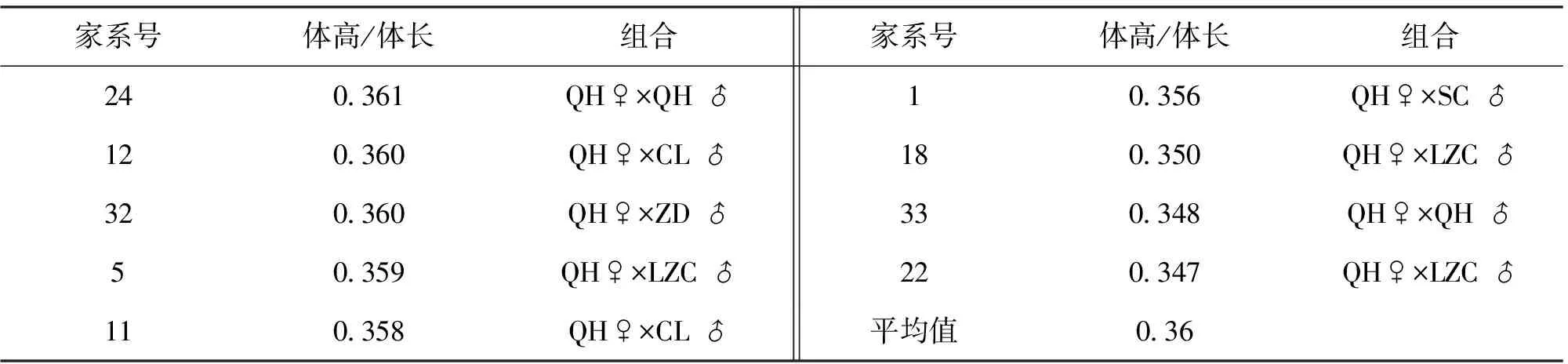
续表5
体高和体质量变异系数最小的都是24号和33号家系,而这2个家系的父本又都是淇河鲫,因此相对于黄河鲤和超级鲤,以淇河鲫作为父本的子代生长性状稳定性更高。

表6 不同家系各性状的变异系数Tab.6 Variable coefficient of body length body height and body weight in deferent families /%
3 讨论
在本实验中,作为同一母本的异精雌核发育家系11号和15号家系无论在养殖存活率(11号家系:42%;15号家系:86%)还是体质量(11号家系:130.81 g;15号家系:96.63 g)方面都存在较大差异,因此异精雌核发育的父本可能在子代的发育过程中起到了一定作用,该结果与之前一些学者在淇河鲫[22]、银鲫(Carassiusauratusgibelio)[23]和彭泽鲫[24]等鲫中的研究结果相同。这可能是由于鲤精子与淇河鲫“杂交”过程中部分异源DNA随机渗入到了淇河鲫的基因组中,也可能是异源精子带入的线粒体、核质等物质影响了子代的某些性状[22],从而产生了异精生物学效应[23]。出现异精生物学效应的原因是由于异源精子的某些遗传物质参与了后代发育,但是不同的异源精子所携带的遗传物质是不一样的,因此对后代的影响也不尽相同,这可能是“同母异父”的11号家系和15号家系在养殖存活率和体质量方面存在显著差异的原因。
淇河鲫不同生长性状的变异系数显示异精雌核发育家系的变异系数要大于淇河鲫群体内建立的家系,表明异源精子的遗传物质不仅影响了雌核发育后代的生长发育,而且对不同个体的影响差异还很大,可能是由于在淇河鲫异精雌核发育过程中出现了异精生物学效应,由于异源精子的遗传物质是随机参与到后代发育过程中的,就造成同一家系后代间也出现较大差异的现象。这与之前研究中认为异育银鲫中的异源精子与银鲫的后代和母本性状一致,而同源精子受精时会产生遗传分化的结果不同[7],至于同源和异源精子在淇河鲫后代中的遗传机理与银鲫有什么区别还需要进一步深入研究。
本研究结果显示,无论是父本半同胞家系还是母本半同胞家系在养殖成活率、生长速度、变异系数和体型等方面均未出现明显的一致性,表明无论采用什么选育方法,父母本对子代均有一定的影响,因此在选育的过程中对父本母本都要进行筛选。另外,根据淇河鲫异精雌核发育后代变异系数较大,而淇河鲫群体内交配变异系数较小的结果(体重变异系数不超过10%),在淇河鲫的选育过程中,可以采用不同异源精子刺激淇河鲫,然后从中筛选出经济性状优良的后代进行雌雄交配,这可作为淇河鲫进行人工选育所采取的育种路线。
参考文献:
[1]王阅雯.淇河鲫鱼形态学和RAPD标记遗传多样性研究[D].河南新乡:河南师范大学,2010.
[2]李学军,刘洋洋.淇河鲫研究进展与开发策略[J].淡水渔业,2012,42(6):93-96.
[3]李学军,王阅雯,高丽霞,等.淇河鲫与野生鲫形态及染色体差异研究[J].水产科学,2010,29(11):674-676.
[4]姚纪花,楼允东.三种群银鲫的RAPD分析初报[J].上海水产大学学报,2000,(1):11-14.
[5] Li F B,Gui J F.Clonal diversity and genealogical relationships of gibel carp in four hatcheries [J].Anim Genet,2008,39(1):28-33.
[6]程 磊,刘 洋,鲁翠云,等.淇河鲫雌核发育克隆系鉴定与性状分析[J].水产学杂志,2013,26(6):1-6.
[7]桂建芳,周 莉.多倍体银鲫克隆多样性和双重生殖方式的遗传基础和育种应用[J].中国科学:生命科学,2010,(2):97-103.
[8] Zhou L,Wang Y,Gui J.Genetic evidence for gonochoristic reproduction in gynogenetic silver crucian carp (CarassiusauratusgibelioBloch) as revealed by RAPD assays [J].J Mol Evol,2000,51(5):498-506.
[9]瞿薇芬,单元勋.河南淇河鲫Carassiusauratus的生物学[J].河南师范大学学报(自然版),1985,(3):53-62.
[10]高春生,范光丽.淇河鲫肌肉营养成分分析及营养价值评定[J].淡水渔业,2006,36(5):33-36.
[11]冯建新,惠 筠,杨兴丽,等.淇河鲫与彭泽鲫的生物学性状比较研究[J].淡水渔业,2004,34(4):27-29.
[12]王德寿.基因组时代养殖鱼类性别可塑性与性控育种研究[C].云南昆明:2015中国遗传学会大会,2015.
[13] Chen S,Zhang G,Shao C,et al.Whole-genome sequence of a flatfish provides insights into ZW sex chromosome evolution and adaptation to a benthic lifestyle [J].Nat Genet,2014,46(3):253-260.
[14] Wu C,Zhang D,Kan M,et al.The draft genome of the large yellow croaker reveals well-developed innate immunity [J].Nat Commun,2014,(5):5227.
[15] Xu P,Zhang X,Wang X,et al.Genome sequence and genetic diversity of the common carp,Cyprinuscarpio[J]Nat Genet.2014,46(11):1212-1219.
[16] Shao C,Bao B,Xie Z,et al.The genome and transcriptome of Japanese flounder provide insights into flatfish asymmetry [J].Nat Genet,2017,49(1):119-124.
[17]李学军,胡灿灿,王 磊,等.鱼类家系选育的研究进展[J].水产科学,2016,(1):81-86.
[18]马爱军,郭建丽,王新安,等.大菱鲆选育家系抗鳗弧菌性能[J].中国水产科学,2014,(3):484-493.
[19]王 磊,胡灿灿,陈松林,等.牙鲆家系4个选育性状的测定与分析[J].中国水产科学,2015,(4):620-629.
[20]王兰梅,朱文彬,董在杰,等.福瑞鲤选育家系不同养殖阶段的生长差异分析[J].南方水产科学,2017,(1):43-49.
[21]杨其彬,温为庚,黄建华,等.斑节对虾4个不同群体建立家系的生长及成活[J].南方水产,2010,(3):36-40.
[22]杨太有,彭仁海,陈宏喜,等.异精雌核发育淇鲫胚胎发育过程中同工酶分析[J].河南师范大学学报(自然科学版),2008,(4):132-135.
[23]蒋一珪,梁绍昌,陈本德,等.异源精子在银鲫雌核发育子代中的生物学效应[J].水生生物学集刊,1983,(1):1-13.
[24]赵 俊,崔 淼,陈湘粦.异精效应在雌核发育彭泽鲫胚胎发育中的同工酶证据[J].水生生物学报,2004,28,(1):33-39.
猜你喜欢
中华实用诊断与治疗杂志(2022年1期)2022-08-31
中华实用诊断与治疗杂志(2022年1期)2022-08-31
农业工程学报(2022年5期)2022-06-22
江苏科技报·E教中国(2020年14期)2020-12-14
口腔医学(2020年11期)2020-11-27
中华戏曲(2020年2期)2020-02-12
中南林业科技大学学报(2019年4期)2019-04-08
人大建设(2018年7期)2018-09-19
森林工程(2018年1期)2018-05-14
农村农业农民·A版(2017年4期)2017-04-20
