
不同择伐方式云南松针叶表型多样性比较*
2018-10-23 11:17蔡年辉杨博宁王亚楠汪梦婷许玉兰李根前
西南林业大学学报 2018年5期
蔡年辉 杨博宁 王亚楠 汪梦婷 许玉兰 李根前
(1. 西南林业大学西南山地森林资源保育与利用教育部重点实验室,云南 昆明 650224; 2. 西南林业大学云南省高校林木遗传改良与繁育重点实验室,云南 昆明 650224)
云南松 (Pinusyunnanensis) 是我国云贵高原的主要乡土树种和重要用材树种[1-3],分布于东经96°~108°、北纬23°~30°之间,其中云南省是云南松的集中分布区[3-5],具有适应性强、耐干旱瘠薄、喜光、木材用途广等特点,是分布区域内瘠薄荒山造林的先锋树种和治理水土流失的重要树种[3,6-7],对分布区域的林业生产和生态经济建设具有举足轻重的作用[3]。但是,目前云南松林分出现稳定性差、遗传品质退化、林分生产力下降等问题[8],如何提高云南松人工林的生态功能和生产功能,并使其得以协调发展和可持续利用是云南林业急需解决的重要问题之一[9],而维持群体遗传多样性显得尤其重要。择伐是森林生态系统最常见的干扰因素之一[10],而不同择伐强度和方式则可能强化了离散性过程,不同择伐方式保留的种质不同,可能对次生种群的遗传结构产生不同影响,进而影响其稳定性[10-11],但不同择伐方式对云南松群体遗传多样性的变化等尚缺乏研究证据。
表型变异反映植物对于环境变化的适应性[12],经过长期的自然选择,性状逐步稳定形成新表型,表型变异在适应进化过程中具有重要的意义[13-15]。研究植物表型多样性,不仅有助于了解环境因子对物种的影响,同时还可以揭示遗传因素与自然因素的关系[16]。同时,表型分析是研究物种遗传多样性的传统方法,可以直观展现群体的遗传结构[17]。叶片是植物光合作用和呼吸作用的主要器官,在植物进化过程中对环境变化比较敏感且可塑性较大,环境变化常导致叶性状的变异以适应不同的环境条件[18-21]。因此,研究植物叶表型多样性在一定程度下可揭示其遗传多样性,前人已对云南松针叶在不同茎干类型及多种环境因子下的多样性特征进行了研究[22-24]。因此,研究不同择伐方式对云南松针叶表型变异的影响,有助于制定更为合理经营措施,为云南松林分的可持续经营提供参考依据。
1 材料与方法
1.1 材料采集
在全面踏查的基础上,根据代表性和典型性原则在云南省双江县 “留优去劣”、“伐优留劣” 和 “无人为择伐” 3种择伐方式下云南松群体设置3个30 m × 30 m样地,3个样地均为云南松主伐后天然更新林分,林分年龄基本一致。2015年11月进行调查和采样,采集样地中每株云南松树冠中上部位2年生针叶,用自封袋保存并进行单株编号,带回实验室备用,每个样地的植株数100~200株。
1.2 性状测定
在所采集的枝条上随机抽取健康完整10束针叶用于表型测定,针叶长、叶鞘长用直尺测量,精确到0.1 cm;针叶粗、针叶束粗用游标卡尺测量,精确到0.01 mm;计算针叶长/针叶粗、针叶长/叶鞘长和针叶束粗/针叶粗3个针叶表型性状指标。
1.3 统计分析
采用SAS 9.0软件对各性状进行巢式方差分析,线形模型为:
Yikj=μ+Si+T(i)j+ε(ij)k
(1)
式中:Yikj为第i个群体第j个家系第k个观测值,μ为总平均值,Si为群体效应 (固定),T(i)j为群体内家系效应 (随机);ε(ij)k为试验误差,分析云南松群体的表型变异特征,并计算表型分化系数[25],计算公式为:
(2)

采用Bio-Dap软件计算针叶性状多样性Shannon-Wiener指数:
H=-∑(Pi)(lnPi)
(3)
式中:Pi为某性状第i个级别出现的概率。
对数据采用Excel软件进行统计,运用SPSS 19.0进行方差分析、相关性分析,并计算平均值、标准差以及变异系数。
2 结果与分析
2.1 云南松针叶表型性状差异
对不同择伐方式下的群体各针叶性状进行方差分析,结果如表1所示。
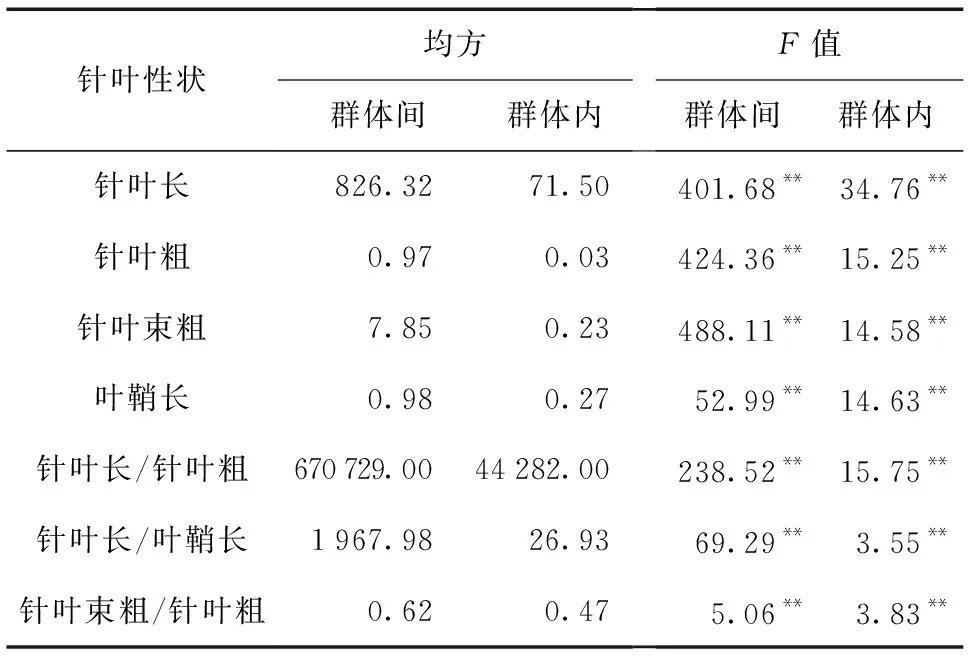
表1 不同择伐方式下云南松针叶表型性状的方差分析Table 1 Variance analysis of needles phenotypic traits of P.yunnanensis in different selective cutting ways
注:**表示差异极显著(P< 0.01)。
由表1可知,各针叶性状在群体间和群体内均存在极显著差异 (P< 0.01),表明云南松针叶性状变异丰富。
进一步对3种择伐方式下针叶表型性状分析结果 (表2) 表明,针叶长、针叶束粗/针叶粗和针叶长/叶鞘长3个表型性状均以无人为择伐方式的稍低,其次为留优去劣方式,而以伐优留劣方式的较高。此外,针叶粗和针叶束粗也以伐优留劣方式的较高。

表2 不同择伐方式下云南松针叶表型性状分析Table 2 Needles morphological traits of P.yunnanensis in different selective cutting ways
注:同列字母不相同表示差异显著 (P< 0.05)。
2.2 云南松针叶性状变异分析
对云南松3个群体叶的形态性状的变异程度进行分析 (表3),云南松7个针叶性状的变异系数范围在14.28%~22.04%,平均变异系数为17.83%,其中针叶长/叶鞘长的变异系数最大为22.04%,叶鞘长次之,为20.08%,针叶长/针叶粗的变异系数也大于平均变异系数,针叶束粗/针叶粗的变异系数最小为14.28%,其他3个性状均小于平均变异系数17.83%。此外,针叶形状指数的变异系数比其单个性状稍大,平均变异系数最大的针叶长/叶鞘长(22.04%)和最小的针叶束粗/针叶粗(14.28%)均为形状指数,说明云南松针叶形状指数比单个性状变异度更大。变异系数间接反映群体的多样性丰富程度,变异系数越大表明性状多样性越丰富。通过对3个群体针叶表型性状变异系数的均值比较可知,3个群体所有性状的平均变异系数相差较小,说明云南松针叶的变异类型分布较均匀,以无人为择伐方式的相对稳定。
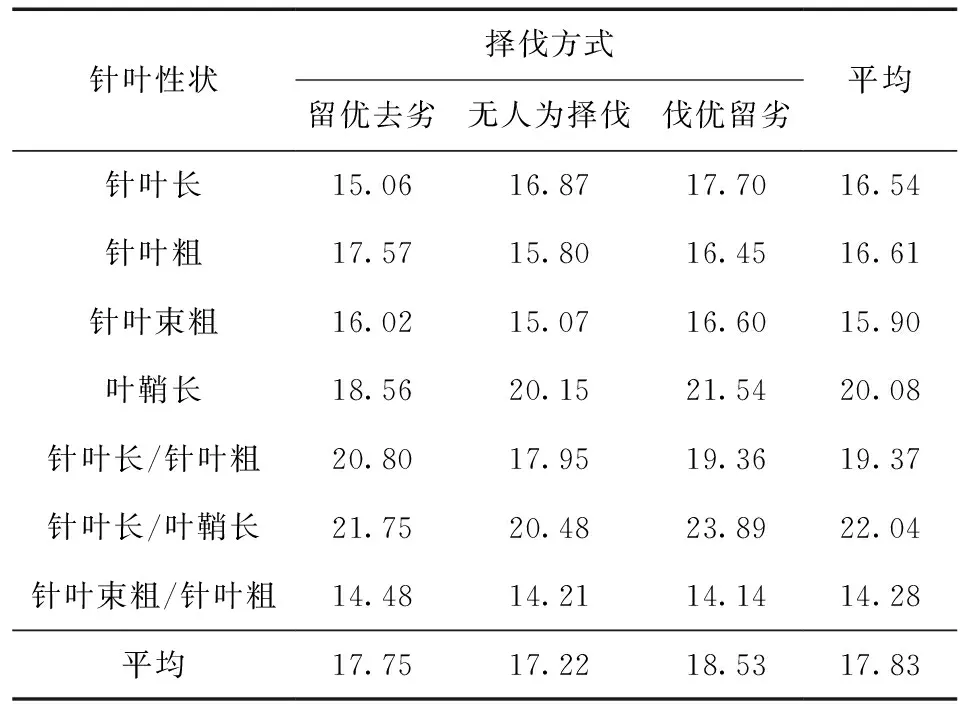
表3 不同择伐方式下云南松针叶表型性状变异系数Table 3 Variation coefficients of needles phenotypic traits of P.yunnanensis in different selective cutting ways %
2.3 云南松针叶表型性状分化
不同择伐方式下云南松针叶表型性状方差分量分析见表4。
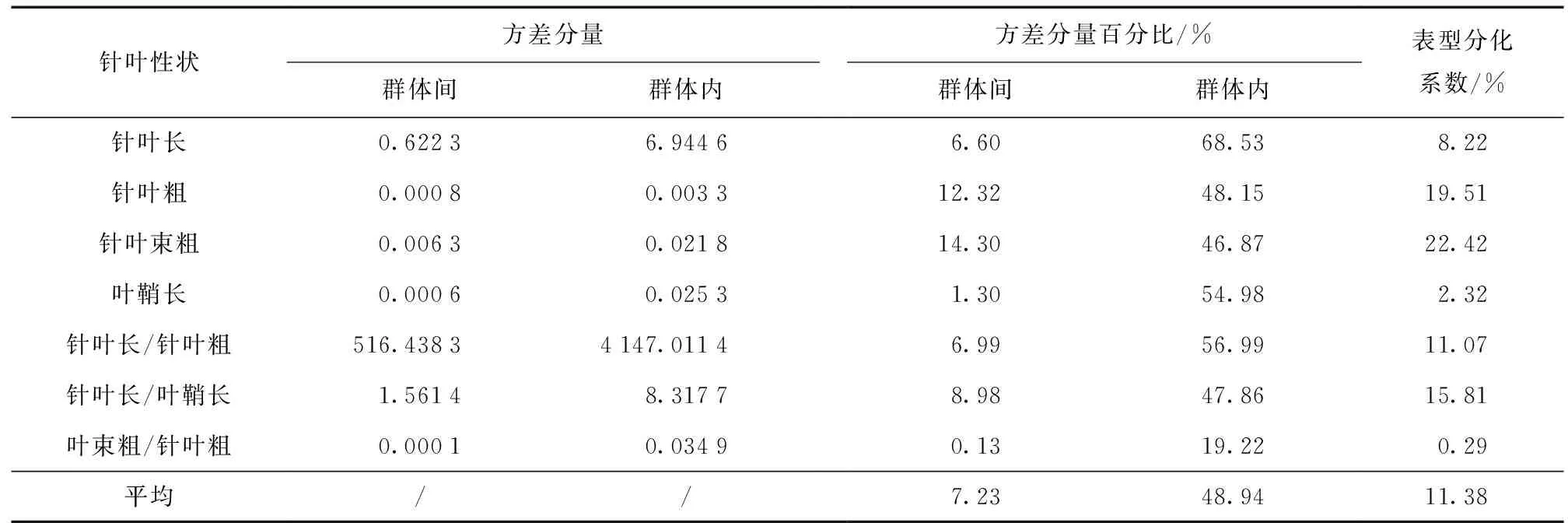
表4 不同择伐方式下云南松针叶表型性状方差分量分析Table 4 Variance components of P.yunnanensis in different selective cutting ways
对3个云南松群体的7个针叶性状进行巢式方差分析,计算出各分量占总变异的比例。由表4可知:云南松7个针叶性状在群体间的平均方差分量为7.23%、群体内为48.94%,即群体间的变异远小于群体内。各表型分化系数幅度为0.29%~22.42%,较大的为针叶粗度和针叶束粗,分别为22.42%和19.51%,最小的为针叶束粗/针叶粗,仅0.29%。7个性状群体间平均表型分化系数为11.38%,即群体间变异占遗传总变异的11.38%,相应地群体内变异占遗传总变异的88.62%,群体间的变异远小于群体内变异,说明群体内变异是云南松针叶性状的主要变异来源。
2.4 云南松针叶表型性状多样性指数
在前述分析的基础上,进一步对各群体的多样性指数进行比较 (表5),结果表明,云南松7个针叶性状的Shannon-Wiener指数变化于1.414 7~1.467 2,各性状的多样性指数差异较小,性状间分布较均匀。3个群体间Shannon-Wiener指数的范围1.405 6~1.457 2,群体间多样性指数差异也较小,与变异系数结果一致,其中以无人为择伐方式的Shannon-Wiener指数略高,以伐优留劣方式的稍低。

表5 不同择伐方式下云南松针叶表型多样性指数Table 5 Shannon-Wiener index of P.yunnanensis in different selective cutting ways
3 结论与讨论
表型多样性是遗传物质多样性的具体表现,表型变异大可能存在的遗传变异大[26-27],反映了群体基因型与环境互作的复杂性、以及适应环境选择压的广泛程度[13],变异的丰富性是群体分化的源泉,也是不同环境选择的结果[28]。云南松针叶表型性状指标在群体间和群体内都存在着显著差异,且群体间的变异小于群体内的变异,表明云南松群体基因型与环境互作的复杂性。云南松分布于东经96°~108°、北纬23°~30°之间,约纵跨8个纬度、横跨12个经度,分布范围广,生境复杂多样[4,29]。植物的变异与自身的遗传特性及生长环境密切相关,一般情况下树种分布范围越大则变异大[30]。由此可以说明,云南松针叶性状变异较大,与分布范围较大相吻合。云南松7个针叶性状变异系数变化波动于14.28%~22.04%,平均17.83%,变化幅度较大,表明云南松针叶表型性状离散程度较高,存在丰富的变异。研究还表明,云南松针叶叶鞘长的变异系数相比其他性状较大,叶鞘是针叶的保护部位,在复杂的生境下,叶鞘响应外界环境的变化而发生改变,从而引起较大的表型变化。此外,不同择伐方式下的表型变异系数均值比较接近,以无人为择伐方式的稍低,即无人为择伐对云南松针叶性状的变化影响更小。在7个针叶表型性状中,针叶长、针叶长/叶鞘长、针叶束粗/针叶粗、针叶粗和针叶束粗5个表型性状均以伐优留劣方式的较高,即在伐优留下方式下,其中针叶长、针叶粗和针叶束粗的生长均有所提高,相应的针叶长/针叶粗、针叶束粗/针叶粗也增加。与其他采伐方式相比,伐优留劣方式是伐去林分中的优势木,对群落的结构、光照、种内竞争等因素的改变较大,环境因素的变化导致针叶表型性状随之变化。因此,生产上应尽量降低人为干扰强度,保持针叶表型性状的稳定。
云南松群体间平均表型分化系数为11.38%,群体间的变异远小于群体内,群体内变异是云南松针叶变异的主要来源。云南松为风媒异花授粉植物,可远距离扩散,从而导致群体间的基因流较大,削弱群体间的遗传分化,从而使群体间的变异降低[31]。云南松群体间平均表型分化系数远低于虫媒花植物山杏 (Prunusarmeniaca) (61.56%)[32]、五角枫 (48.42%)[33]和葛萝槭 (Acergrosseri) (50.16%)[34],与风媒花授粉植物西加云杉 (Piceasitchenrsis) (10.1%)[35]、马尾松 (Pinusmassoniana) (6.44%)[25]和云南红豆杉 (Taxusyunnanensis) (14.66%)[36]大致相同,大多数风媒花植物其群体间表型分化系数要低于虫媒花植物[13]。与此同时,低的表型分化系数表明3种不同择伐方式对群体遗传结构的影响较小。
云南松不同择伐方式群体间的Shannon-Wiener′s多样性指数差异较小,其中以无人为择伐方式的稍高,一方面说明不同择伐方式对云南松群体遗传多样性影响不大,另一方面表明无人为择伐对遗传多样性有一定的维持作用。有研究报道认为择伐或其他经营活动会引起种群密度下降或生境片段化,从而引起种群等位基因的丢失和遗传多样性的下降[37],异交率下降,近交率上升[38],改变种群的遗传结构[39]。Wang等[40]以云杉 (Piceaasperata) 未破坏的天然林、择伐后种植与择伐后天然更新的成熟林和幼林为对象,研究显示天然成熟林的遗传多样性最高,而择伐后种植和天然更新的成熟林或幼林的遗传多样性均没有显著差异,择伐后幼林和未受破坏过的天然成熟林间遗传相似性高、遗传分化低,这也说明未受破坏过的天然成熟林起着遗传多样性储藏库的作用。由此可见,林木择伐后引起遗传多样性的改变,与林木自身特性有关,如果树种的种子和花粉传播能力强,则对遗传多样性无显著影响。
猜你喜欢
翠苑(2019年4期)2019-11-11
种子(2019年5期)2019-07-02
农村百事通(2019年8期)2019-05-13
作文周刊·小学四年级版(2019年8期)2019-04-23
浙江中医杂志(2019年5期)2019-01-06
国际木业(2018年3期)2018-09-10
农机化研究(2018年2期)2018-06-05
农业环境科学学报(2017年11期)2017-12-07
南方农业·下旬(2017年8期)2017-10-23
江苏农业科学(2016年6期)2016-07-25
