
千里光菲灵碱诱导MEK/ERK1/2介导的宫颈癌细胞自噬效应
2018-10-26 06:54马景蕃叶甘萍韦启良邱龙新
中国药科大学学报 2018年5期
马景蕃,张 燕,叶甘萍,韦启良,邱龙新*
(1龙岩学院生命科学学院,龙岩 364012;2预防兽医学与生物技术福建省高校重点实验室,龙岩 364012;3福建省家畜疫病防治与生物技术重点实验室,龙岩 364012)
在世界范围内,宫颈癌严重威胁着广大育龄妇女的健康和生命,它是妇女第二常见的恶性肿瘤,同时也是妇女发病和死亡的主要原因之一,近几年其发病率呈现年轻化趋势[1]。目前,化疗是宫颈癌患者主要治疗手段之一,但在临床上化疗产生的耐药使得宫颈癌治疗失效和复发,如何克服宫颈癌化疗耐药是其治疗面临的一大挑战[2]。
自噬是真核生物细胞内普遍存在的一种现象,诸多研究表明[3],自噬是一把双刃剑:一方面,细胞通过自噬可以实现代谢需要和某些细胞器的更新,起到细胞保护作用;另一方面,自噬也可以诱导细胞发生自噬性死亡。随着人们对自噬作用及其调控机制的深入研究,很多证据表明[4],自噬在肿瘤的发生发展及肿瘤治疗中起着重要作用。
千里光菲灵碱(seneciphylline,SENE)是一种来源于菊科植物[Gynurasegetum(Lour) Merr]根部的生物碱[5],近年来,国内外学者对千里光菲灵碱的抗肿瘤作用较为关注,研究表明千里光菲灵碱能抑制多种肿瘤细胞的增殖[6],但其抗肿瘤的作用机制尚不明确,有待进一步研究。本研究采用荧光共定位、Western blot等研究方法,观察千里光菲灵碱对宫颈癌HeLa、Caski细胞的影响,并探讨其可能机制,为其进一步的临床应用提供理论依据。
1 材 料
1.1 药物与试剂
DMEM高糖培养基、胎牛血清均(美国Hyclone 公司);千里光菲灵碱[成都德思特生物技术有限公司,用二甲基亚砜(DMSO)溶解];MTT试剂盒、ECL化学发光试剂(上海碧云天生物技术有限公司);Lipofectamine®3000、Lysotraker Red(美国Invitrogen公司);海藻糖(trehalose,Tre)、3-甲基腺嘌呤(3-methyladenine,3MA)、氯喹(chloroquine,CQ)、U0126(上海Sigma-Aldrich公司)。LC3抗体(美国Novus公司);GAPDH抗体(美国Millipore Chemicon公司);p62抗体(美国Santa Cruz公司);P-ERKl/2(Thr202/Thr 204)及T-ERKl/2抗体(美国Cell Signaling Technology公司);其他试剂均为市售分析纯。
1.2 仪 器
酶标仪Elx-800型(美国Bake公司);CFM-550倒置荧光显微镜(德国Zeiss公司);Versa MAXTM多功能酶标仪(美国Molecular Devices公司);CO2培养箱(美国Thermo公司)。
1.3 细胞株
人宫颈癌HeLa、Caski细胞株购于ATCC,用含10%的胎牛血清、青霉素和链霉素(各100 U/mL)的DMEM培养液在37℃,5%CO2的饱和湿度条件下进行传代培养,当细胞处于对数生长期时用于实验。
1.4 动 物
25只雄性BALB/c裸鼠购于上海斯莱克实验动物公司,许可证号SCXK(沪)2017-0005,体质量18~20 g。
2 方 法
2.1 MTT法检测细胞增殖
将HeLa、Caski细胞分别接种于96孔板中,调整细胞密度为每毫升6×104个细胞,培养24 h后,分对照组与实验组进行实验,对照组不加千里光菲灵碱,实验组用浓度为25、50、100、200、400、800、1 000 μmol/L的千里光菲灵碱分别处理细胞24、48、72 h,每组设6个重复。培养结束后加入5 mg/mL MTT液20 μL,在37 ℃避光培养箱孵育4 h后吸去上清液,再每孔中加入DMSO 120 μL,摇床轻微振荡15 min,用酶标仪于570 nm处测量各孔的吸收度,并计算细胞存活率。细胞存活率(%)=(药物组吸收度/对照组吸收度)×100。
2.2 细胞转染及荧光显微镜观察
根据Lipofectamine®3000转染试剂盒说明书,将GFP-LC3质粒转染体系分别转入到HeLa、Caski细胞中,使用G418筛选出具有稳定转染的HeLa、Caski细胞。将生长状态良好的GFP-LC3/HeLa、GFP-LC3/Caski细胞接种到24孔板中,接种密度为每孔3×105个细胞,培养20 h后,对细胞进行相应实验处理,处理结束后在倒置荧光显微镜下观察GFP聚集的产生情况并进行图片采集。
2.3 Lysotraker Red染色
用100 mol/L海藻糖(Tre)和250 μmol/L千里光菲灵碱分别处理GFP-LC3/HeLa、GFP-LC3/Caski细胞24 h后,加入75 nmol/L的 Lysotraker Red染色25 min,弃去上清液,经PBS洗涤后,倒置荧光显微镜观察染色情况并拍照。
2.4 Western blot分析
将对数生长期的HeLa、Caski细胞接种于6孔板中,处理结束后,用RIPA裂解液裂解细胞,提取总蛋白,各组取蛋白样品30 μg上样,进行SDS-PAGE电泳,将蛋白转移到PVDF膜上,于室温下封闭1.5 h,加入一抗4 ℃孵育过夜,TBST缓冲液洗涤3次,二抗室温摇床孵育2 h,TBST缓冲液洗涤3次,ECL化学发光试剂显影。
2.5 裸鼠皮下移植瘤模型建立及抑制实验
将对数生长期的HeLa、Caski细胞以每只1×107个细胞的注射量分别接种于裸鼠右侧腋窝皮下。当瘤体积达200 mm3时进行体内抑瘤实验。将25只荷瘤鼠均分为5组:对照组(生理盐水组)、HeLa细胞低剂量组(1 mg/kg)、HeLa细胞高剂量组(5 mg/kg)、Caski细胞低剂量组(1 mg/kg)、Caski细胞高剂量组(5 mg/kg)。隔日灌胃,共14 d。每日观察荷瘤裸鼠的一般状况及皮下移植瘤的生长状态。瘤体积(mm3)=1/2长×宽2,抑制率(%)=(Vc-Vt)/Vc×100,其中Vc为对照组平均瘤体积,Vt为各实验组平均瘤体积。
2.6 统计分析
3 结 果
3.1 千里光菲灵碱对HeLa、Caski细胞增殖的影响
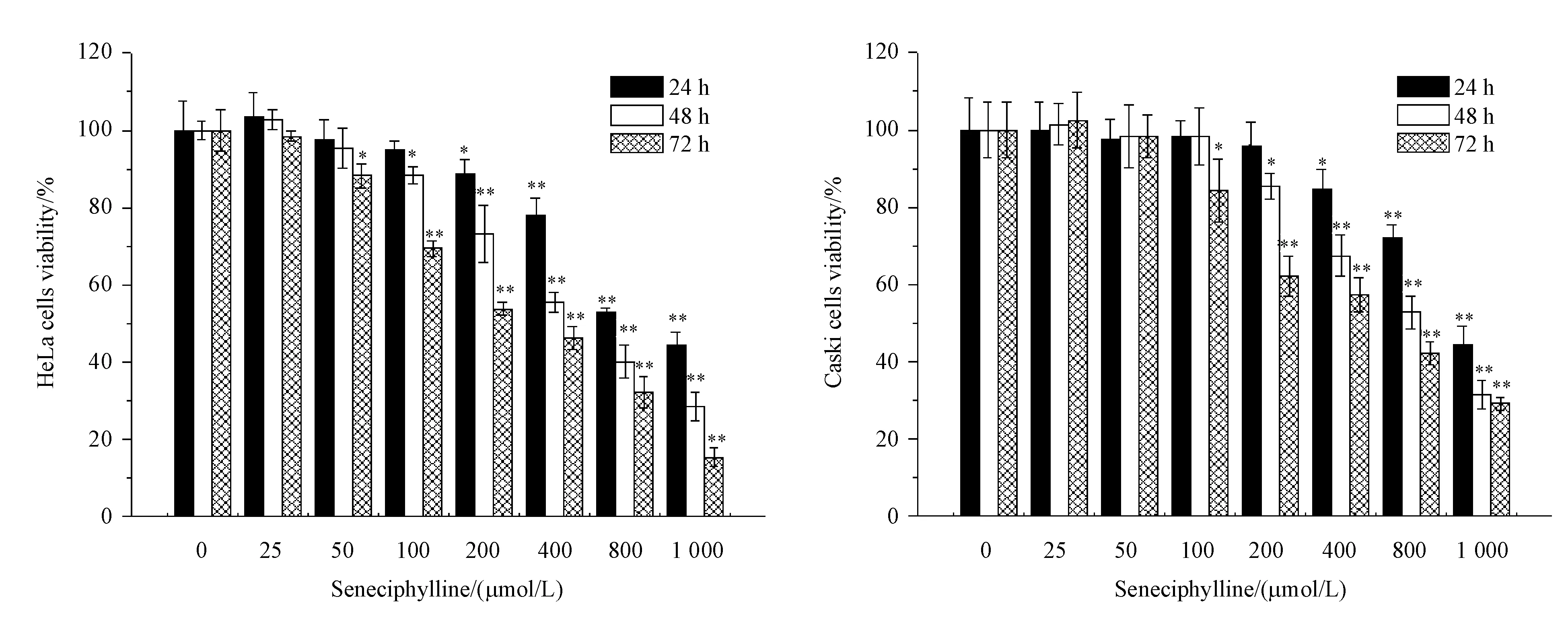
图1结果显示,不同浓度的千里光菲灵碱分别处理HeLa、Caski细胞24、48、72 h后,千里光菲灵碱对两种宫颈癌细胞增殖抑制作用均呈时间、剂量依赖性。图1-A显示,当处理时间为24、48、72 h,千里光菲灵碱浓度分别为200、100、50 μmol/L时HeLa细胞存活率与对照组均呈显著性差异(P<0.05)。当HeLa细胞培养24、48、72 h的IC50分别为882.8、494.2、330.6 μmol/L;图1-B显示,当处理时间为24、48、72 h,千里光菲灵碱浓度分别为400、200、100 μmol/L时Caski细胞存活率与对照组均呈显著性差异(P<0.05)。Caski细胞培养24、48、72 h的IC50分别为923.3、646.5、457.6 μmol/L。
3.2 千里光菲灵碱诱导宫颈癌细胞发生自噬
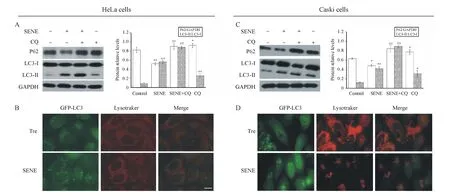
GFP-LC3/HeLa、GFP-LC3/Caski是稳定转染并持续表达绿色荧光标签的人宫颈癌细胞系,当自噬被激活后,GFP-LC3-I就会被加工成GFP-LC3-II,聚集在自噬小体膜中,在荧光显微镜下,能够观测到具有绿色荧光的点状聚集(自噬体)。图2-A,D显示,分别处理GFP-LC3/HeLa、GFP-LC3/Caski细胞24 h后,对照DMSO组均不能引起两种人宫颈癌细胞中出现GFP的点状聚集,而经典的自噬诱导剂海藻糖(Tre)组出现了明显的GFP点状聚集;用不同浓度的千里光菲灵碱处理两种细胞24 h后,出现的绿色荧光点状聚集均呈现剂量依赖效应,即随千里光菲灵碱处理浓度的增加,绿色荧光点状聚集阳性细胞的比例增多。以上结果表明千里光菲灵碱可能诱导了HeLa及Caski细胞发生自噬。
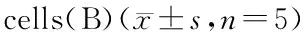
*P<0.05,**P<0.01vscontrol group
为了进一步证实此结果,用Western blot分析了自噬相关蛋白(LC3和P62)的表达情况。如图2-B所示,不同浓度的千里光菲灵碱处理HeLa细胞24 h后,经药物处理(200、400、800 μmol/L)的HeLa细胞与对照(0 μmol/L)相比,其P62/GAPDH比值显著降低,且呈浓度依赖性,而LC3-II/LC3-I比值显著提高,亦呈浓度依赖性;200 μmol/L的药物处理Caski细胞24 h后,其P62/GAPDH比值及LC3-II/LC3-I比值与对照(0 μmol/L)相比无显著性差异,而400和800 μmol/L的药物处理Caski细胞24 h后,其P62/GAPDH比值与对照(0 μmol/L)相比显著降低,且呈浓度依赖性,而LC3-II/LC3-I比值显著提高,亦呈浓度依赖性(图2-E)。另外,400 μmol/L的千里光菲灵碱分别处理HeLa及Caski细胞不同时间后,P62/GAPDH比值和LC3-II/LC3-I比值与药物处理呈时间依赖性(图2-C,F)。以上结果均表明千里光菲灵碱能诱导宫颈癌HeLa及Caski细胞发生自噬。
3.3 千里光菲灵碱诱导宫颈癌细胞产生完整自噬流
将千里光菲灵碱(400 μmol/L)与饱和浓度的自噬的下游抑制剂氯喹(CQ,50 μmol/L)单独或联合处理HeLa及Caski细胞24 h后,与对照组相比,氯喹单独处理组在两种细胞中均出现了显著的P62及LC3-II的累积,说明氯喹阻断了HeLa及Caski细胞自噬下游底物的降解。与单独千里光菲灵碱处理组相比,千里光菲灵碱联合氯喹处理组在两种细胞中具有显著的LC3-II及P62的蛋白累积(图3-A,C),这说明千里光菲灵碱并没有阻断自噬下游底物的降解。
随后,进一步用荧光共定位方法进行验证,Lysotraker Red是一种细胞碱性染料,它可以选择性地给细胞内部的溶酶体染色,同时亦可指示溶酶体的合成及活性[7]。如图3-B,D显示,用100 mmol/L海藻糖和400 μmol/L千里光菲灵碱分别处理GFP-LC3/HeLa和GFP-LC3/Caski细胞24 h,千里光菲灵碱与阳性对照海藻糖一样,均能够诱导HeLa 和Caski细胞内出现绿色荧光点聚集(自噬体),且这些荧光点聚集可与Lysotraker Red染色的溶酶体实现共定位。这进一步说明了千里光菲灵碱不会干扰自噬体与溶酶体的融合。
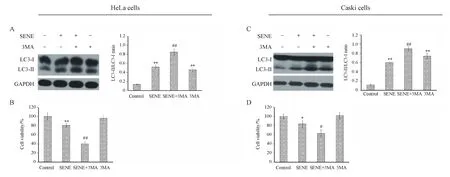
3.4 抑制自噬增强千里光菲灵碱对宫颈癌细胞增殖的抑制作用
自噬对肿瘤细胞的增殖具有双重作用,为明确千里光菲灵碱诱导的自噬与宫颈癌细胞增殖的关系,将自噬抑制剂与千里光菲灵碱联合作用于HeLa和Caski细胞。3-甲基腺嘌呤(3MA)是一种通过抑制Ⅲ型PI3K复合物的自噬上游抑制剂[8]。将千里光菲灵碱(400 μmol/L)与自噬抑制剂3MA(5 mmol/L)单独或联合处理HeLa及Caski细胞24 h后,与对照组相比,3MA单独处理组在两种细胞中均出现了显著的LC3-II的累积,说明3MA阻止了HeLa和Caski细胞自噬的发生。与单独千里光菲灵碱处理组相比,千里光菲灵碱联合3MA处理组在两种细胞中均具有更高的LC3-II/LC3-I比值,呈极显著性差异(图4-A,C)。两种细胞活力检测(图4-B,D)表明,与单独千里光菲灵碱处理组相比,千里光菲灵碱联合3MA处理组显著抑制了两种细胞的增殖。

A:Green fluorescence puncta in HeLa cells;B&C:Autophagy related protein expression level in HeLa cells;D:Green fluorescence puncta in Caski cells;E&F:Autophagy related protein expression level in Caski cells
Tre:Trehalose (100 mmol/L);SENE:Seneciphylline.Scale bar=20 μm
*P<0.05,**P<0.01vscontrol group
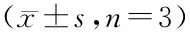
A:Autophagy related protein expression level in HeLa cells;B:Co-localizations between the GFP-LC3 dots and Lysotracker Red in HeLa cells;C:Autophagy related protein expression level in Caski cells;D:Co-localizations between the GFP-LC3 dots and Lysotracker Red in Caski cells
Scale bar=20 μm
*P<0.05,**P<0.01vscontrol group;#P<0.05,##P<0.01vsSENE group
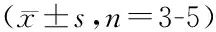
A:Autophagy related protein expression level in HeLa cells;B:HeLa cells viability;C:Autophagy related protein expression level in Caski cells;D:Caski cells viability
*P<0.05,**P<0.01vscontrol group;#P<0.05,##P<0.01vsSENE group
3.5 千里光菲灵碱诱导的宫颈癌细胞自噬依赖于MEK/ERK1/2途径
为明确千里光菲灵碱诱导的自噬是否激活MEK/ERK1/2途径,进行Western bolt实验和免疫荧光检测。U0126和PD98059是MEK特异性的磷酸化抑制剂,而MEK的活性抑制将直接导致ERK的活性抑制[9]。将千里光菲灵碱(400 μmol/L)与U0126(10 mmol/L)/PD98059(50 mmol/L)单独或联合处理HeLa或Caski细胞24 h后,与对照组相比,单独千里光菲灵碱处理组在两种细胞中均出现了显著的P-ERK1/2蛋白累积,即千里光菲灵碱诱导了ERK的活化。而与单独千里光菲灵碱处理组相比,千里光菲灵碱联合U0126/ PD98059处理组在两种宫颈癌细胞中均抑制了千里光菲灵碱诱导的ERK活化,两组呈极显著性差异,而HeLa及Caski细胞内部总ERK蛋白含量没有明显变化(图5-A,B,D,E);同时,图5-C,F表明,U0126/PD98059抑制了千里光菲灵碱在两种细胞中诱导的自噬体形成。综上,千里光菲灵碱诱导的细胞自噬激活了MEK/ERK1/2途径。
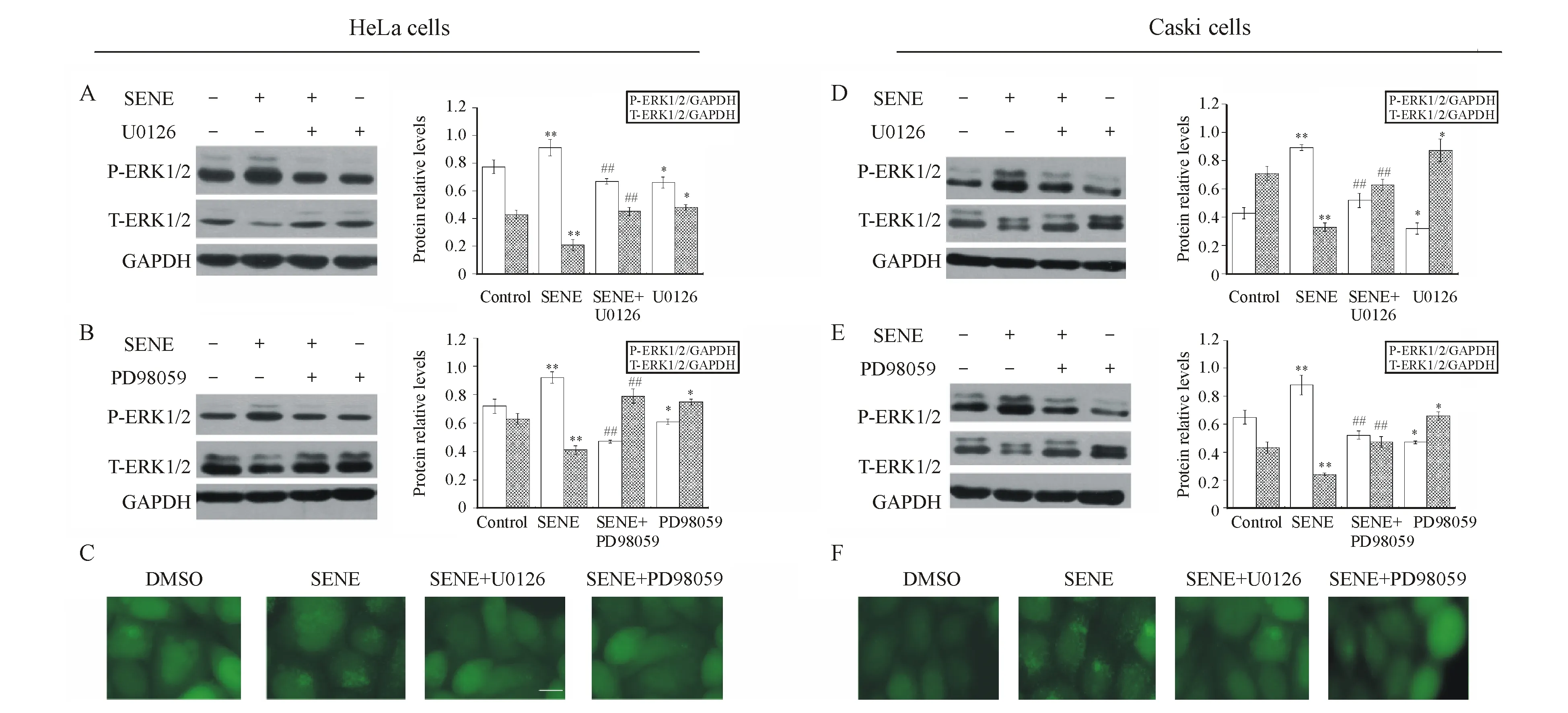
A&B:ERK protein expression level in HeLa cells;C:Green fluorescence puncta in HeLa cells;D&E:ERK protein expression level in Caski cells;
F:Green fluorescence puncta in Caski cells
Scale bar=20 μm
*P<0.05,**P<0.01vscontrol group;#P<0.05,##P<0.01vsSENE group
3.6 千里光菲灵碱对荷瘤鼠移植瘤生长的抑制作用
如图6所示,各处理荷瘤鼠肿瘤体积均显著小于对照组的瘤体积,接种HeLa细胞并灌胃千里光菲灵碱低剂量组(1 mg/kg)的抑瘤率为22.97%,接种HeLa细胞并灌胃千里光菲灵碱高剂量组(5 mg/kg)的抑瘤率为33.22%,接种Caski细胞并灌胃千里光菲灵碱低剂量组(1 mg/kg)的抑瘤率为18.37%,接种Caski细胞并灌胃千里光菲灵碱高剂量组(5 mg/kg)的抑瘤率为27.68%。

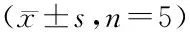
*P<0.05,**P<0.01vscontrol group
4 讨 论
传统中药活性物质的抗肿瘤研究越来越受到研究学者们的关注。本研究表明,千里光菲灵碱可以抑制人宫颈癌HeLa及Caski细胞的增殖,且其抑制作用呈时间、剂量依赖性。因对后续细胞自噬及其机制研究均需细胞具有一定活力,故本研究根据千里光菲灵碱对两种细胞作用24 h的IC50,选取了200、400、800 μmol/L 3个浓度进行实验。
自噬是细胞质内出现的一种自我保护过程,其在维持细胞发育及肿瘤发生发展中发挥着重要作用。大量研究表明,很多抗肿瘤药物如麦冬皂苷B[10]、盐酸青藤碱[11]等可以不同程度地诱导肿瘤细胞发生自噬。Liu等[6]研究表明,千里光菲灵碱可以抑制肿瘤细胞的增殖,但其是否能诱导人宫颈癌细胞发生自噬及其机制尚无报道。当细胞内自噬被激活后,微管相关蛋白LC3-I就会被加工成LC3-II,并最终被溶酶体融合降解,P62蛋白可与微管相关蛋白LC3结合并被降解[12]。因此,二者常被选作细胞发生自噬的标志蛋白。本研究结果显示,千里光菲灵碱处理HeLa及Caski细胞后,出现了明显的GFP点状聚集,另外,Western blot实验表明,千里光菲灵碱可以促进LC3-I向LC3-II蛋白转换,同时,P62蛋白累积减少。这些结果均表明千里光菲灵碱诱导HeLa及Caski细胞发生了自噬。
自噬是一个完整的过程,包括双层分隔膜的产生,双层分隔膜包裹待降解物形成自噬体,自噬体与溶酶体融合,降解自噬底物。当自噬性降解被抑制,如自噬体与溶酶体融合被阻或溶酶体功能受损,均可导致细胞内出现自噬现象。因此,实验中出现的GFP点状聚集、LC3-II蛋白的累积并不能说明细胞是否发生了完整自噬。氯喹是细胞发生自噬的下游抑制剂,主要通过改变溶酶体的pH,阻断自噬体与溶酶体的融合从而阻止自噬底物的降解,饱和浓度的氯喹(50 μmol/L)可以完全抑制自噬底物的降解[13]。本研究的研究结果表明,在饱和浓度的氯喹完全抑制自噬的同时,千里光菲灵碱可进一步促进P62和LC3-II蛋白在HeLa及Caski细胞内的累积。此外,荧光共定位实验表明,千里光菲灵碱诱导的GFP点聚集可与LysotrakerRed染色的溶酶体实现共定位,表明了自噬体与溶酶体融合形成了自噬溶酶体。因此,本研究认为千里光菲灵碱可诱导HeLa及Caski两种宫颈癌细胞发生完整的自噬流。
自噬是一种高度动态的过程,其在肿瘤的发生、发展中具有两面性。为了明确千里光菲灵碱诱导的自噬是导致自噬性死亡还是导致细胞对千里光菲灵碱的耐药性增加,本研究使用了自噬抑制剂3MA,3MA是一种蛋白激酶抑制剂,它主要作用于细胞内的Ⅲ类磷脂酰肌醇-3激酶(PI3K),而PI3K是激活自噬上游一种重要的激酶,最近许多研究者[14]发现其与抗肿瘤药物联合作用可增加药物对肿瘤细胞的敏感性。本研究的研究结果表明,3MA可促进LC3-II蛋白在HeLa及Caski细胞内的累积,同时,千里光菲灵碱联合3MA具有更为显著的LC3-II蛋白累积。MTT细胞增殖实验表明,自噬上游抑制剂3MA提高了千里光菲灵碱对HeLa及Caski细胞增殖的抑制作用。表明自噬导致了宫颈癌细胞对千里光菲灵碱的抗肿瘤作用降低,自噬阻断可使千里光菲灵碱对宫颈癌细胞增殖的抑制作用增加,这与大多数中药活性成分抗肿瘤研究相一致[15],即自噬减弱了抗肿瘤药物的抗肿瘤作用,使得药物的耐药性增加,抑制自噬可以提高药物对肿瘤细胞的敏感性,增加对肿瘤细胞增殖的抑制作用。
细胞外调节蛋白激酶(ERK),可以感知细胞内外的信号刺激,其与细胞的增殖、转分化等现象关系密切[16-17]。众多研究结果显示,ERK信号途径参与对细胞自噬的调节。为了进一步明确千里光菲灵碱诱导宫颈癌细胞发生自噬的分子机制,本研究对MEK/ERK1/2信号途径中的关键蛋白ERK进行了研究。结果表明千里光菲灵碱诱导的HeLa及Caski细胞自噬激活了MEK/ERK1/2途径。另外,抑制自噬提高千里光菲灵碱对HeLa及Caski细胞的抗肿瘤效果是否与促进细胞凋亡有关还需做进一步研究。本研究选用宫颈癌HeLa及Caski细胞裸鼠皮下移植瘤为实验模型,进一步探讨千里光菲灵碱对移植瘤生长的抑制作用。结果表明,千里光菲灵碱的高剂量组(5 mg/kg)的抑瘤效果最好。
综上所述,千里光菲灵碱可以抑制人宫颈癌HeLa、Caski细胞的增殖,并能诱导完整的自噬流,抑制细胞自噬可增强千里光菲灵碱的抗肿瘤作用,千里光菲灵碱诱导的宫颈癌细胞自噬激活了MEK/ERK1/2信号通路,千里光菲灵碱可显著抑制宫颈癌细胞裸鼠皮下移植瘤的生长,这为千里光菲灵碱对人宫颈癌的临床治疗提供了实验依据和理论基础。
猜你喜欢
河南科技(2022年6期)2022-04-22
昆明医科大学学报(2022年3期)2022-04-19
中老年保健(2021年12期)2021-08-24
昆明医科大学学报(2021年8期)2021-08-13
天津医科大学学报(2021年3期)2021-07-21
中华养生保健(2020年10期)2021-01-18
中国生殖健康(2020年7期)2021-01-18
中国比较医学杂志(2020年11期)2020-12-09
读与写(2019年35期)2019-11-05
中央民族大学学报(自然科学版)(2018年3期)2018-11-09
