
蛋白体外结合实验联合质谱分析鉴定Num1的互作蛋白
2018-12-29 00:52唐仙英
中南民族大学学报(自然科学版) 2018年4期
唐仙英,肖 瑶,海 力
(中南民族大学 生命科学学院,武陵山区特色资源植物种质保护与利用湖北省重点实验室,武汉430074)
为确保细胞的命运决定因子准确地分配到每个子细胞,所有真核细胞均须正确定位有丝分裂纺锤体[1].细胞质动力蛋白Dynein在各类细胞的纺锤体定位过程中起关键作用[2-4].在芽殖酵母的不对称分裂过程中,为了能沿胞质微管产生拉力移动纺锤体,Dynein在与微管相连的同时,需被锚定在细胞膜上.Dynein在细胞膜上的锚定依赖于Num1(Nuclear migration 1,核迁移1)[5-7],Num1是迄今为止所发现的参与纺锤体定位的唯一位于细胞膜上的因子.Num1蛋白大约313 kDa,由多个结构域组成.其中,位于其C末端的PH结构域通过与细胞膜上的磷酸肌醇PI(4,5)P2相互作用介导Num1与细胞膜结合[8,9];而另一个位于其N末端含有两段预测的Coiled-coil序列的结构域则通过自我相互作用介导Num1在细胞膜上形成由14个分子组成的补丁状复合物,因此被命名为Patch Assembly(PA)结构域[10].此外,PA结构域也介导Num1与Dynein的相互作用[10].
定位于细胞膜上的Num1复合物不具有运动性[8, 11],除了在纺锤体定位中的作用,它还通过与线粒体膜上特定种类的脂类物质结合而介导线粒体与细胞膜的连接,并由此调节线粒体的分裂[12,13].Lackner 等[10, 12]发现线粒体与细胞膜的连接是由Num1的PA结构域尤其是其中预测的Coiled-coil序列所介导,但Num1在纺锤体定位与线粒体中的活性如何调节目前尚不清楚.本研究利用在BL21大肠杆菌细胞中表达纯化的PA结构域,通过Pull-down实验分离酵母细胞中与PA结构域相互作用的蛋白质,并进一步通过质谱分析完成鉴定,旨在通过鉴定Num1的互作蛋白,为揭示Num1的作用途径及机制奠定基础.
1 材料与方法
1.1 材料和仪器
DH5α和BL21大肠杆菌细胞(北京鼎国昌盛);蛋白酶抑制剂混合物(Protease Inhibitor Cocktail, Roche),S蛋白琼脂糖(S protein agarose slurry)、Bugbuster蛋白提取反应液(Novagen);氨苄青霉素、氯霉素、异丙基硫代半乳糖苷IPTG、苯甲基磺酰氟PMSF(BioSharp).
高速冷冻离心机(CR22G型, HITACHI);微量冷冻离心机(FC5515R型, OHAUS);高压细胞破碎仪(BT40/TS2/AA型, Constant System Cell Disruptor);凝胶成像分析仪(Universal Hood II型, BIO-RAD).
1.2 质粒与酵母菌株
编码Num1PA(1-303)-PCN-S-TEV-Z的质粒pXT65(PCN: PreScission蛋白酶酶切位点,S: S-tag, TEV: TEV蛋白酶酶切位点,Z: IgG binding motif)用于表达PA结构域与S-tag的融合蛋白,其构建方法如下:编码Num1 1-303 aa且在5′端含有一个NcoI限制性内切酶位点,3′端含有一个NotI限制性内切酶位点的DNA片段分别用正向引物5′-CCAGCCATGGCCTCCCACAACAACAGGCATAAAAAG-3′和反向引物5′-CCAGGCGGCCGCCAGATGTTACTGTAGTATCG-3′从酵母基因组DNA通过PCR扩增,扩增的片段经NcoI和NotI酶切后与同样经NcoI和NotI酶切的载体pBSG01[10]连接并经测序确认,产生pXT65.pXT66(PCN-S-TEV-Z)为空载体对照.酵母菌株YWL555[10].
基因型为MATαnum1Δ∷HIS3ura3-52lys2-801leu2-Δ1his3-Δ200trp1-Δ63.
1.3 大肠杆菌细胞培养
质粒制备或转化用DH5α细胞,重组蛋白表达和制备用BL21细胞[14].DH5α细胞用含氨苄青霉素的LB培养基于37 ℃培养.用于制备重组蛋白的BL21细胞按下述方式培养:保存于-80 ℃的BL21大肠杆菌细胞在含氨苄青霉素的LB固体培养基上37 ℃过夜培养.次日从LB平板上挑取细胞接种到含氨苄青霉素和氯霉素的LB液体培养基,37 ℃过夜培养.第3 d将液体培养基中的细胞稀释至OD600=0.1,25 ℃培养3 h,加入异丙基硫代半乳糖苷(IPTG)至终浓度为0.5 mmol/L,20 ℃培养16 h诱导融合蛋白表达,离心收集细胞.
1.4 蛋白可溶性检测
按1.2培养的BL21细胞经离心收集后,按每0.01 g细胞加入50 μL Bugbuster蛋白提取反应液,混匀,室温下轻摇15 min,4 ℃ 16000 g 离心20 min;分别取上清和沉淀加入蛋白上样缓冲液,沸水煮5 min.将上清和沉淀制备的蛋白样品分别经变性聚丙烯酰胺凝胶电泳(SDS-PAGE)分离后用考马斯亮兰染色,比较目的蛋白在上清和沉淀中的分配比例.
1.5 Pull-down实验
1.5.1 重组蛋白的制备
携带pXT65和pXT66的BL21大肠杆菌按1.3各收集5 g细胞.按照每克细胞1 mL的比例加入Bind/Wash buffer [150 mmol/L NaCl, 20 mmol/L Tris-Cl (pH 7.5), 0.05% TritonX-100, 1 mmol/L EDTA, 1 mmol/L DTT, Protease Inhibitor Cocktail],用高压细胞破碎仪(压力207 MPa, 4 ℃)破碎细胞.破碎的细胞4 ℃下12000 g 离心10 min,收集上清,此为大肠杆菌细胞裂解液.取100 μL S protein agrose悬液,1000 g离心1 min,除去上清,用Bind/Wash buffer洗3次,每次1 mL.加入大肠杆菌细胞裂解液,4 ℃旋转混合45 min,离心,除去上清.再用Bind/Wash buffer洗珠子3次,每次5 mL,洗过的珠子置于冰上.
1.5.2 酵母细胞裂解液的制备
离心收集YPD液体培养基中旺盛生长的对数期YWL555细胞,每个pull-down实验需要5 g湿细胞. 按1 mL/g的比例加入酵母裂解缓冲液[30 mmol/L HEPES (pH 7.4), 50 mmol/L乙酸钾, 2 mmol/L醋酸镁, 0.2 mmol/L EGTA, 0.05% Triton X-100, 1 mmol/L DTT, Protease Inhibitor Cocktail (Roche)],用高压细胞破碎仪破碎细胞.破碎的细胞液10000 g离心5 min,上清即为酵母细胞裂解液.
1.5.3 Pull-down反应
将1.5.2中制备的酵母细胞裂解液加入到1.5.1中已结合了重组蛋白的S蛋白琼脂糖中,4 ℃旋转2 h,1000 g离心1 min,移去上清.用Wash buffer [10 mmol/L Tris-Cl (pH 8.0), 150 mmol/L KCl, 10% Glycerol, 0.05% Triton X-100, 1 mmol/L DTT, Protease Inhibitor Cocktail (Roche)] 洗3次,每次8 mL,除去Wash buffer,加入100 μL 蛋白上样缓冲液,沸水煮5 min,收集上清保存于-20℃.
1.6 SDS-聚丙烯酰胺凝胶电泳及蛋白鉴定
所有的蛋白质样品均用10%变性聚丙烯酰胺凝胶电泳进行检测[15].其中Pull-down沉淀下来的酵母蛋白样品10% SDS-聚丙烯酰胺凝胶电泳分离后,再用考马斯亮蓝染色液染色,切下被PA结构域融合蛋白沉淀下来的特异性蛋白条带进行鸟枪法(shotgun)LC-MS质谱分析.
2 结果与分析
2.1 PA结构域重组蛋白的表达
PA结构域是预测的Coiled-coil结构,高度不可溶.比较了不同培养条件下Num1PA(1-303)-PCN-S-TEV-Z融合蛋白在BL21大肠杆菌中的表达(见图1),最后确定适宜的诱导条件为IPTG浓度0.5 mmol/L,20 ℃过夜培养,此时Num1PA(1-303)-PCN-S-TEV-Z存在于细胞裂解液上清的比例相对较高,图1中虚线框对应预期蛋白带,箭头指示为该蛋白在20 ℃条件下表达(图1b)较37 ℃下表达(图1a)时存在于裂解液上清中的比例高.

1,3,5,7)上清;2,4,6,8) 沉淀a) 37 ℃, IPTG 0.5 mmol/L; b) 20 ℃, IPTG 0.5 mmol/L图1 不同条件下蛋白诱导表达结果Fig.1 Protein expression induced under different conditions
2.2 PA结构域重组蛋白的纯化
携带pXT65[Num1PA(1-303)-PCN-S-TEV-Z]或pXT66(PCN-S-TEV-Z)的BL21大肠杆菌细胞在含0.5 mmol/L IPTG浓度的培养基中20 ℃过夜培养诱导蛋白表达.由于诱导表达的Num1PA(1-303)-PCN-S-TEV-Z蛋白含有一个S-tag,因此可在裂解大肠杆菌细胞后,使其与S蛋白琼脂糖珠子结合而得到纯化,结果见图2.如图2所示:S蛋白琼脂糖能从表达pXT65的大肠杆菌细胞裂解液中沉淀下来Num1PA(1-303)-PCN-S-TEV-Z蛋白(箭头所示),而未从表达空载体pXT66的大肠杆菌细胞裂解液中沉淀下来蛋白.
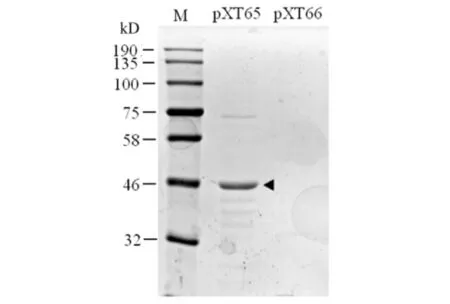
图2 Num1PA(1-303)-PCN-S-TEV-Z蛋白的纯化Fig.2 Purification of Num1PA(1-303)-PCN-S-TEV-Z protein
2.3 PA结构域重组蛋白与酵母蛋白的离体结合
为分离与PA结构域相互作用的酵母蛋白,用结合在S蛋白琼脂糖珠子上的Num1PA(1-303)-PCN-S-TEV-Z蛋白与酵母细胞裂解液进行了离体结合实验,即Pull-down实验.由于PA结构域具有自我相互结合的特性[10],为防止PA结构域与酵母内源的Num1蛋白相结合,酵母细胞裂解液从已敲除了野生型NUM1基因的酵母菌株(num1Δ,即YWL555)中制备.洗去非特异性结合的蛋白质,即获得了结合在S蛋白琼脂糖珠子上的Num1PA(1-303)-PCN-S-TEV-Z及与其结合的其他蛋白质的复合物.此蛋白复合物样品经SDS-PAGE分离后用考马斯亮蓝染色,结果见图3.如图3所示:上箭头为Num1PA(1-303)-PCN-S-TEV-Z蛋白,下箭头是被Num1PA(1-303)-PCN-S-TEV-Z特异性沉淀下来的蛋白复合物(即pXT66的Pull-down样品中缺乏的蛋白),将其切割下来进行质谱分析,用于鉴定与PA结构域互作的蛋白.

图3 Num1PA(1-303)-PCN-S-TEV-Z的Pull-down蛋白检测
Fig.3 Detection of proteins pulled down by Num1PA(1-303)-PCN-S-TEV-Z
2.4 互作蛋白的鉴定
通过对Pull-down沉淀下来的蛋白复合物进行鸟枪法(shotgun)LC-MS质谱分析,鉴定出一系列新的与Num1相互作用的蛋白,按照参与的细胞功能分组如表1.
3 结语
位于Num1 N末端的PA结构域(1-303 aa)在Num1的功能中起核心作用,缺失了PA结构域的Num1将同时失去其在纺锤体定位和线粒体中的功能[10,12,13].为分离Num1的互作蛋白,在大肠杆菌细胞中表达了PA结构域和纯化标签的融合蛋白Num1PA(1-303)-PCN-S-TEV-Z,并用S protein agarose对其进行了纯化.结合在S protein agarose珠子上的重组蛋白用于从不表达内源Num1蛋白的酵母细胞裂解液中沉淀PA结构域的互作蛋白.通过对沉淀下来的蛋白进行鸟枪法LC-MS质谱分析,鉴定了一系列新的Num1的互作蛋白,包括大量的核糖体蛋白和参与蛋白折叠转运的蛋白,参与基因转录和翻译的核酸酶和蛋白酶,及其他功能各异的蛋白质.这些结果为进一步揭示Num1在纺锤体定位及线粒体中的作用机制奠定了基础,也为此过程的调控机制提供了更多思路.

表1 Num1的互作蛋白Tab.1 Proteins interacting with Num1
猜你喜欢
九江学院学报(自然科学版)(2022年2期)2022-07-02
食品与发酵工业(2022年9期)2022-05-17
热带海洋学报(2022年2期)2022-03-31
太原理工大学学报(2021年6期)2021-11-25
心肺血管病杂志(2020年5期)2021-01-14
中成药(2019年12期)2020-01-04
读与写(2019年31期)2019-11-26
中国海洋大学学报(自然科学版)(2019年7期)2019-05-21
中国海洋大学学报(自然科学版)(2019年7期)2019-01-04
药学研究(2015年11期)2015-12-19

- 中南民族大学学报(自然科学版)的其它文章
- 催化剂Ni-CeO2的制备及其在甲烷二氧化碳重整反应中的催化性能
- 脊髓灰质炎病毒I型单克隆抗体的制备
- 浅析丛枝菌根共生的生态学意义
- 以碱性聚合酶2(PB2)为靶点的抗流感病毒药物的虚拟筛选
- Discovery of Potential Novel TRPC 4 Inhibitor by Combination of Virtual Screening and 3D-QSAR Modeling
- Optical and Electrical Properties of Organic Semiconductor Thin Films for Optoelectronic Devices