
藏北高山嵩草草甸群落特征及生产力对模拟增温幅度的响应
2019-02-20 10:43李军祥张扬建朱军涛常文静刘瑶杰俎佳星朱艺旋陈宁
生态学报 2019年2期
李军祥,张扬建,朱军涛,曾 辉,常文静,丛 楠,刘瑶杰,6,俎佳星,6,黄 珂,6,朱艺旋,6,王 荔,唐 泽,6,陈宁,6
1 北京大学深圳研究生院,深圳 518055 2 拉萨高原生态试验站,生态系统网络观测与模拟重点实验室,中国科学院地理科学与资源研究所,北京 100101 3 中国科学院青藏高原地球科学卓越创新中心,北京 100101 4 中国科学院大学资源与环境学院,北京 100190 5 北京大学城市与环境学院,北京 100871 6 中国科学院大学,北京 100190
全球气候变暖已经是毋庸置疑的事实[1],尤其是青藏高原表面的温度上升要高于全球平均气温的上升速率[2- 3]。气候变化与植物群落特征及生态系统初级生产力的变化有着密切的联系[4- 6],尤其在两极及高寒地区表现为最强[7]。植物群落特征及生态系统生产力的变化对地球表面生态系统的结构和功能[8- 9]及对人类服务功能的可持续发展极其重要[10],因此,在气候变化背景下,研究植物群落特征及生态系统生产力变化的机制对应对未来气候变化具有重要的意义[11- 12]。
在全球变暖背景下,有大量通过田间试验或模型模拟陆地生态系统植物群落特征及植被地上生产力对气候变化响应的研究,但不同物种与功能型植物及植被地上生产力对气候变暖的响应规律尚不统一[5- 6,13- 16]。研究显示增温提高了植被生产力[13- 14],或抑制植被生产力[7,15],或没有改变植被生产力[16];不同物种或功能型之间的响应存在差异[6,17],如增温抑制了禾草和莎草的生长[17],或促进了禾草和莎草的生长[18]。全球变暖是如何影响植物群落特征及地上生产力,目前尚不清楚[12- 15]。理清温度升高对植物群落特征及地上生产力的影响机制,对准确地评估生态系统生产力具有非常重要的意义[1,10]。
增温对植物群落特征及地上生产力的影响主要取决于不同植物物种或功能群的生长[5- 9]。而影响植物生长的因素众多,如植物群落组成[19- 20]、物种的不同步性[1]、植物根系分布深度、植物物候、微生物群落的组成[21]、植物养分的分配[22]等间接的影响了植物的生长[13- 14,23]。此外,增温对植物生长的影响是受特殊生态系统环境及增温对非生物特性影响的限制,如增温减少土壤水分抑制了植被生产力的累积[24]。增温对植物生长的影响还取决于植物本身的特性,如相比抗旱能力差的植物,抗旱能力好的植物可以减少增温引发的干旱胁迫的限制[25],同时增温促进豆科植物氮的固定,可能会增加土壤速效氮,促进植被生物量的生产[26- 27]。
青藏高原气候严酷,陆地表层生态系统脆弱,其生态系统功能和结构对气候变化的响应极其敏感[1,28],然而到目前为止,有关温度升高对青藏高原腹地高寒草甸植物群落特征及地上生产力的研究,大多数只检验增温和不增温的效果[1,5],而模拟不同增温梯度对高寒草甸植物生长和植物群落组成影响的研究较少。因此,本研究利用开顶箱(OTCs)式设置不同增温梯度实验(W1、W2、W3、W4),研究不同增温梯度对藏北高寒草甸植物群落特征及地上生产力的影响。其目的是在探究全球变暖的背景下,增温对高寒草甸植物群落特征及地上生产力的影响,探明影响高寒草甸植物群落特征及地上生产力的关键环境要素,为青藏高原高寒草甸生态系统应对未来气候变化提供基础数据和理论依据。
1 材料与方法
1.1 试验区概况
本研究区位于青藏高原腹地的藏北高原草地生态系统研究站,简称那曲站(31°38.513′N, 92°0.921′E),海拔大约4600 m。那曲站是隶属于中国科学院地理科学与资源研究所的拉萨高原生态实验站,位于藏北羌塘高原核心地带,那曲地区那曲县那曲镇曲果仁村境内,距那曲县城约22 km。该地区是典型的亚寒带气候区的高寒草甸草原,年平均气温-0.9℃至-3.3℃,全年日照时数2788 h;年平均降水量431.7 mm,降水主要集中在6月至9月,占年降水量的85%。高寒缺氧,全年气候干冷,无绝对的无霜期,昼夜温差大,每年10月至次年5月为风雪期和土壤冻结期,植物生长季一般从6月到9月。该地区降水来源主要是夏季季风,一般在6月底或7月初到来,且表现出很大的年际差异[29]。
植被类型为高寒草甸草原,植被覆盖度为60%—90%,以高山嵩草(Kobresiapygmaea)为主,其他伴生种包括早熟禾(Poaannua)、黄花棘豆(Oxytropisochrocephala)、钉柱委陵菜 (Potentillasaundersiana)、楔叶委陵菜 (P.cuneata)、二裂委陵菜 (P.bifurca)、紫花针茅(Stipapurpurea)、无茎黄鹌菜 (Youngiasimulatrix)、藏西风毛菊 (Saussureaalpine)等,土壤类型为高寒草甸土[30]。
1.2 试验设计
本实验研究对象为自然状态下的高寒草甸生态系统,采用开顶箱(OTCs)式增温装置。利用OTCs的高度控制增温的效果,设置了4个增温梯度(W1、W2、W3、W4),见图1。如表1,每个OTCs的顶边长均为80 cm,保证所有处理的OTCs开口大小一致,底边长分别为100、110、120、130 cm,高度分别为40、60、80、100 cm。OTCs均采用三角钢支撑,四周材质为透明玻璃纤维,透光率达到90%以上。因此,本实验设置5种处理,包括对照(C),增温1 (W1)、增温2 (W2)、增温3 (W3)、增温4 (W4),其中每个处理设置4个重复,共计20个样方。OTCs于2013年9月在那曲站完成安装[30]。

表1 OTCs的规格/cm
W1:增温1,Warming 1;W2:增温2,Warming 2;W3:增温3,Warming 3;W4:增温4,Warming 4

图1 OTCs 示意图Fig.1 Open top chambers (OTCs) used in the study
1.3 群落及生物量的测定
在不同增温实验处理样地中心布设一个50 cm×50 cm的固定样方,在每年生长季利用点框架法(即:每个样方的每个物种都覆盖的基础上用100个(5 cm×5 cm)均匀分布的网格估计)经行群落特征的估测,包括每个物种的盖度、高度、多度等,以及群落的总盖度和平均高度等。并在8月中旬(生长旺盛期)在每个样方内取10 cm×10 cm的面积齐地面剪草,收割地上生物量,在65℃下烘干至恒重并称重。同时,为了避免OTCs的边缘效应,只在OTCs的中心区域内取样。
1.4 环境因子的观测
利用维萨拉公司的 HMP155A 探头(Vaisala, Helsinki, Finland)每天记录OTCs内地上10 cm的空气温度,且探头外部用防辐射罩保护。利用坎贝尔公司的 CS655 探头(Campbell Scientific, Logan, UT)每天记录表层5 cm的土壤温湿度。每个试验处理均安装2套空气和土壤温湿度探头,小气候数据为2套探头的平均值。
1.5 数据处理
在高山嵩草草甸生态系统中,高山嵩草为优势种,委陵菜属植物为伴生种,其他物种植物为罕见种,且高山嵩草和委陵菜属植物在群落中所占的比重大于85%,因此,本研究将高山嵩草草甸群落划分为高山嵩草、委陵菜属植物和其他物种植物等三个植物功能群。物种的重要值采用如下公式:重要值=(相对盖度+相对频度+相对高度+相对密度)×100/4。
统计分析采用双因素方差分析(Two-way ANOVA with Turkey′s test),分别检验增温处理、年际作用及其交互作用对地上生物量、群落总盖度、高山嵩草盖度、委陵菜属植物盖度和其他物种总盖度的影响,差异显著性水平设定为0.05。采用单因素方差分析,检验增温处理对地上生物量、群落总盖度、高山嵩草盖度、委陵菜属植物盖度和其他物种总盖度的影响,差异显著性水平设定为0.05。线性回归分析土壤温度、土壤水分与地上生物量、群落总盖度、高山嵩草盖度、委陵菜属植物盖度和其他物种总盖度之间的关系。以上分析在 SPSS version 18.0 中进行。
2 结果
2.1 微环境变化
2015年整个生长季的降水量为263 mm(DOY:150—DOY:250),低于多年平均降水量389 mm,生长季的平均气温是9.77℃。2016年整个生长季的降水量为412 mm(DOY:150—DOY:250),高于多年平均降水量389 mm,生长季的平均气温是9.86℃。在藏北高原OTCs增温方式提高了空气和土壤温度,而降低了土壤水分 (图2)。在2015年,与对照相比,实验增温(W1、W2、W3、W4)分别提高了空气温度2.5、3.0、3.6、4.1℃,分别提高了土壤温度0.3、1.4、1.9、2.3℃;增温 (W1、W2、W3、W4)分别降低了土壤水分2.5%、4.7%、5.8%、7.3%。在2016年,与对照相比,实验增温 (W1、W2、W3、W4) 分别提高了空气温度1.7、2.3、2.9、3.5℃,分别提高了土壤温度0.3、1.4、1.9、2.1℃;增温 (W1、W2、W3、W4) 分别降低了土壤水分4.2%、7.6%、10.7%、12.4%。
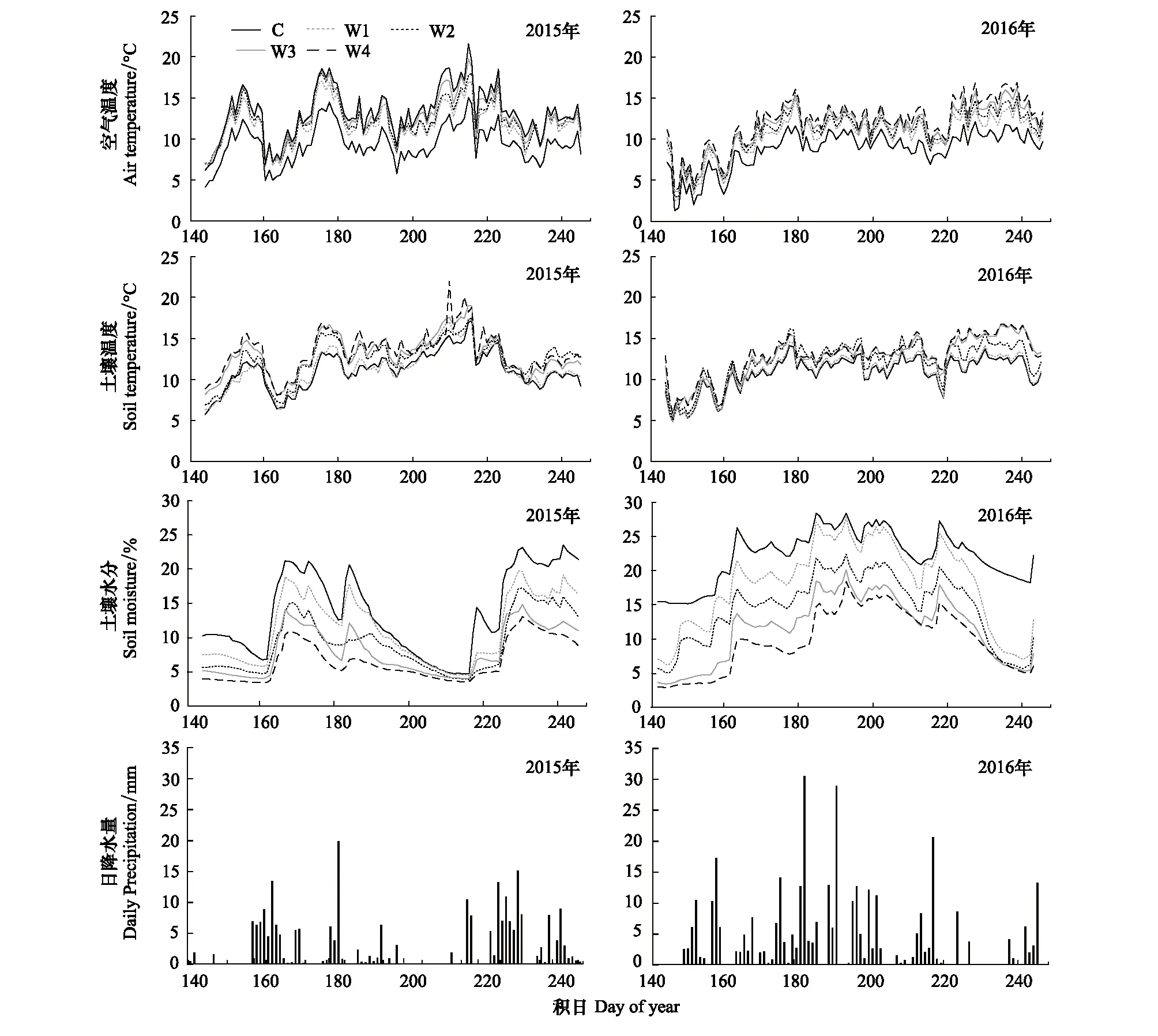
图2 增温对空气温度、土壤温度、土壤水分的影响及生长季日降水量Fig.2 Soil temperature, air temperature, soil moisture under an experimental warming gradient, and daily precipitationC:对照,Control;W1:增温1,Warming 1;W2:增温2,Warming 2;W3:增温3,Warming 3;W4:增温4,Warming 4
2.2 地上生物量的变化
如表2所示,增温显著影响了群落的地上生物量(P<0.001),且地上生物量存在显著的年际间差异(P<0.05)。与对照相比,在2015年,增温(W1、W3、W4)减少了地上生物量分别为7%(P>0.05)、40%(P>0.05)、69%(P<0.05),而增温(W2)增加了地上生物量7%(P>0.05);在2016年,增温(W1、W3、W4)减少了地上生物量分别为3%(P>0.05)、49%(P>0.05)、82%(P<0.05) (图3),而增温(W2)增加了地上生物量13%(P>0.05) (图3),说明低度增温对植物地上生物量的累积无影响,与之相反,高度增温抑制了植物地上生物量的累积。在不同实验处理下,对照和增温(W1、W2、W3)处理的地上生物量在2016年分别高于2015年26%、37%、40%、8%(P>0.05,图3),而增温(W4)在2016年低于2015年27%(P>0.05,图3)。

表2 增温对地上生物量、群落总盖度及植物功能群盖度的影响(F值)
W:增温,Warming;Y:年,Year;***,**,*分别表示P<0.001,P<0.01,P<0.05的差异性
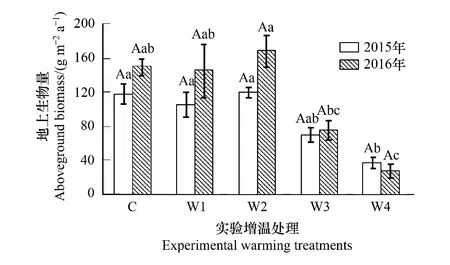
图3 不同幅度增温对地上生物量的影响 Fig.3 Responses of aboveground biomass to a warming gradient不同大写字母表示在相同增温处理下,2015年与2016年差异显著 (P<0.05),不同小写字母表示同一年不同增温处理间差异显著 (P<0.05),相同字母之间不显著
2.3 增温对植物地上群落的影响
如表3所示,不同幅度增温样地与对照样地的群落物种组成存在明显差异。在2015年,对照样地共有12个物种,增温处理(W1、W2、W3、W4)样地分别是12、11、9、8个物种;在2016年,对照样地共有11个物种,增温处理(W1、W2、W3、W4)样地分别是10、9、7、6个物种。在2015年生长季,与对照相比,增温处理(W1、W3、W4)少了物种蒲公英(Taraxacummongolicum),增温处理(W2、W3、W4)少了雪灵芝(ArenariakansuensisMaxim)和附地菜(Trigonotispeduncularis),且增温处理(W4)少了火绒草(Leontopodiumcalocephalum),与之相反,增温处理(W1、W2)多了物种黄芪(Astragaluspulvinatus);而在2016年生长季,与对照相比,增温处理(W1、W2、W3、W4)少了物种蒲公英(T.mongolicum)和雪灵芝(A.kansuensisMaxim),增温处理(W3、W4)少了物种附地菜(T.peduncularis)和藏西凤毛菊(S.alpine),且增温处理(W4)少了物种紫花针茅(S.purpurea)和无茎黄鹌菜(Youngiasimulatrix),与之相反,增温处理(W1、W4)多了物种黄芪(Astragaluspulvinatus)。
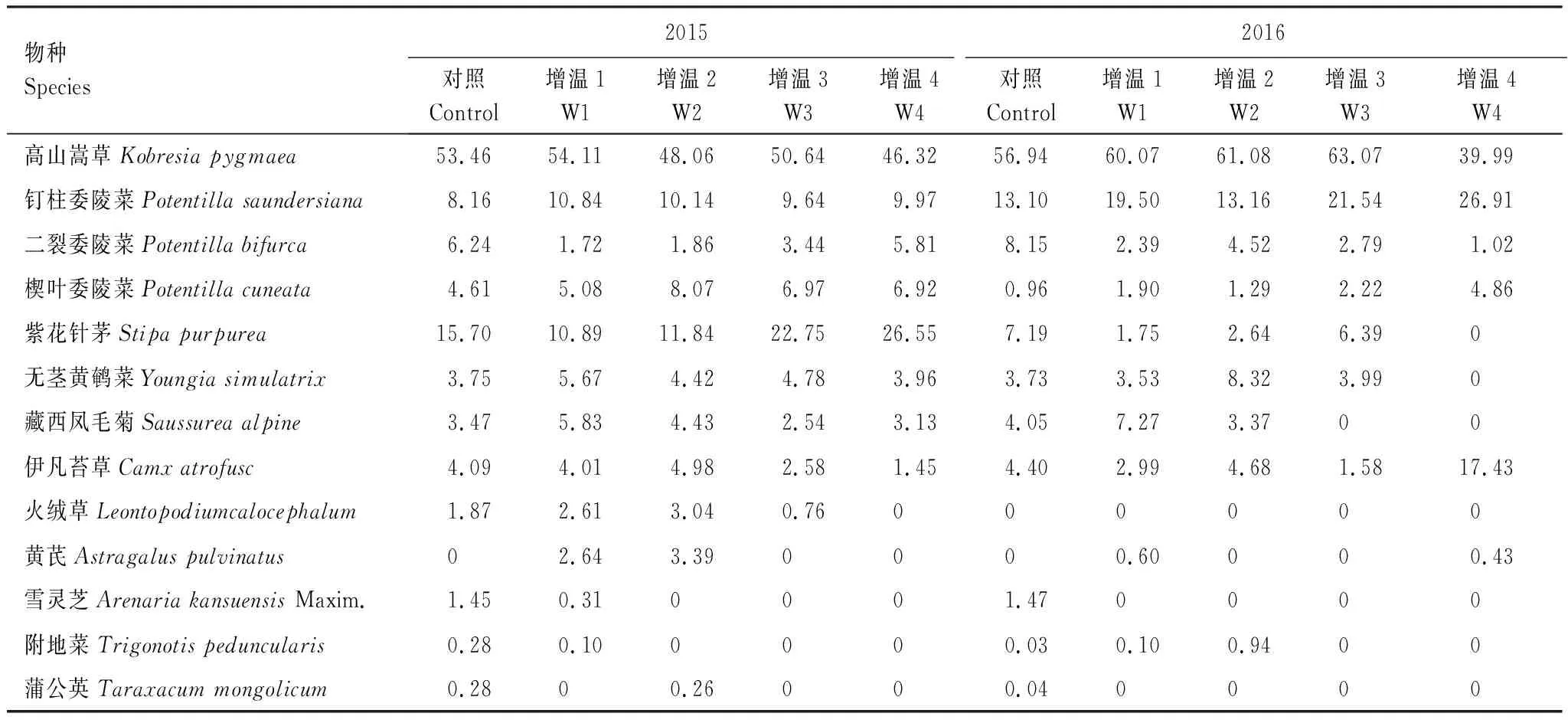
表3 不同幅度增温对高寒草甸植物群落的重要值的影响
不同幅度增温样地与对照样地的群落物种的重要值存在明显差异。比较所有样地内共同出现物种的重要值,发现高山嵩草和钉柱委陵菜(P.saundersiana)在所有群落中占有绝对优势,且增温增加了钉柱委陵菜的重要值。增温减少了二裂委陵菜(P.bifurca)的重要值,与之相反,增温增加了楔叶委陵菜(P.cuneate)的重要值。在2015年生长季,增温(W1)样地内的物种雪灵芝重要值下降最为显著(P<0.05),增温(W4)样地内的物种高山嵩草重要值下降最为显著(P<0.05),与之相反,增温(W3、W4)样地内的物种紫花针茅重要值增加最为显著(P<0.05)。在对照样地内,由高山嵩草和紫花针茅占优势的群落物种组成,经不同幅度增温后,逐渐演变为由高山嵩草和紫花针茅占绝对优势的群落物种组成。在2016年生长季,增温(W1)样地内的物种二裂委陵菜重要值下降最为显著(P<0.05),增温(W3、W4)样地内的物种二裂委陵菜重要值下降最为显著(P<0.05),增温(W4)样地内的物种高山嵩草重要值下降最为显著(P<0.05),与之相反,增温(W2)样地内的物种无茎黄鹌菜重要值增加最为显著(P<0.05),增温(W4)样地内的物种钉柱委陵菜重要值增加最为显著(P<0.05)。在对照样地内,由高山嵩草和钉柱委陵菜占据绝对优势的群落物种组成,经不同幅度增温后,逐渐演变为由高山嵩草、钉柱委陵菜和伊凡苔草(Camxatrofusc)占优势的群落物种组成。
如表2所示,增温显著改变了群落总盖度、高山嵩草盖度和委陵菜属植物盖度(P<0.05),且群落总盖度、高山嵩草盖度和其他物种总盖度存在显著地年间差异(P<0.05),增温和年际间交互作用显著地影响了群落总盖度和其他物种盖度(P<0.05)。在2015年,与对照相比,增温(W1、W2、W3、W4)显著减少了群落总盖度和高山嵩草盖度分别为28%、23%、59%、60%和26%、33%、61%、64%(P<0.01,图4),增温(W3、W4)显著减少了委陵菜属植物盖度分别为58%、60%(P<0.05,图4),与之相反,增温(W1、W2、W3、W4)对其他物种总盖度均无显著影响(P>0.05,图4),且增温(W1、W2)对委陵菜属植物盖度均无显著影响(P>0.05,图4)。在2016年,与对照相比,增温(W1、W2、W3)对群落总盖度、高山嵩草盖度和委陵菜属植物盖度均无显著影响(P>0.05图4),而增温(W4)分别显著减少了83%、85%和71%(P<0.01图4)。同时增温(W1、W2)对其他物种总盖度均无显著影响(P>0.05,图4),而增温(W3、W4)显著减少了其他物种总盖度分别为80%、96%(P<0.05,图4)。
不同幅度增温对群落总盖度、高山嵩草盖度和其他物种盖度的影响在也存在年际差异(图4)。在不同实验处理下,对照和增温(W1)处理的群落总盖度在2016年分别显著高于2015年46%、64%(P<0.01,P<0.05,图4),而增温(W2、W3、W4)无显著差异;对照和增温(W1、W2、W3)处理的高山嵩草盖度在2016年分别显著高于2015年112%、118%、124%、261%(P<0.05,图4),而增温(W4)无显著差异;增温(W1、W2、W3、W4)处理的其他物种总盖度在2016年分别显著低于2015年73%、67%、77%、96%(P<0.05,图4),而对照无显著差异。

图4 不同幅度增温对群落总盖度及其组分的影响Fig.4 Responses of community coverage and composites to a warming gradient不同大写字母表示在相同增温处理下,2015年与2016年差异显著 (P<0.05),不同小写字母表示同一年不同增温处理间差异显著 (P<0.05),相同字母之间不显著
2.4 影响地上生产力的因子
线性回归分析表明,在2015年和2016年地上生物量与土壤水分呈显著正相关关系(P<0.001,r2=0.54, 0.51,图5),而与土壤温度呈显著负相关关系(P<0.001,r2=0.49, 0.43,图5),说明随着土壤温度的增加,地上生物量累计减少,而土壤水分越高,地上生物量累计的越多。在2015年和2016年,群落总盖度、高山嵩草盖度和委陵菜属盖度与土壤水分呈显著正相关关系(全部P<0.05,r2= 0.80, 0.49, 0.85, 0.32, 0.42, 0.34,图5),与土壤温度呈显著负相关关系(全部P<0.05,r2=0.71, 0.34, 0.81, 0.22, 0.39, 0.32,图5),说明土壤水分越高越利于高山嵩草和委陵菜属的生长,与之相反,土壤温度越低越利于高山嵩草和委陵菜属的生长。在2015年和2016年,其他物种总盖度与土壤温度没有明显的相关性(P>0.05,图5),而与土壤水分呈显著正相关关系(P=0.02, 0.03,r2=0.28, 0.32,图5),说明土壤水分越高越利于其他物种的生长。
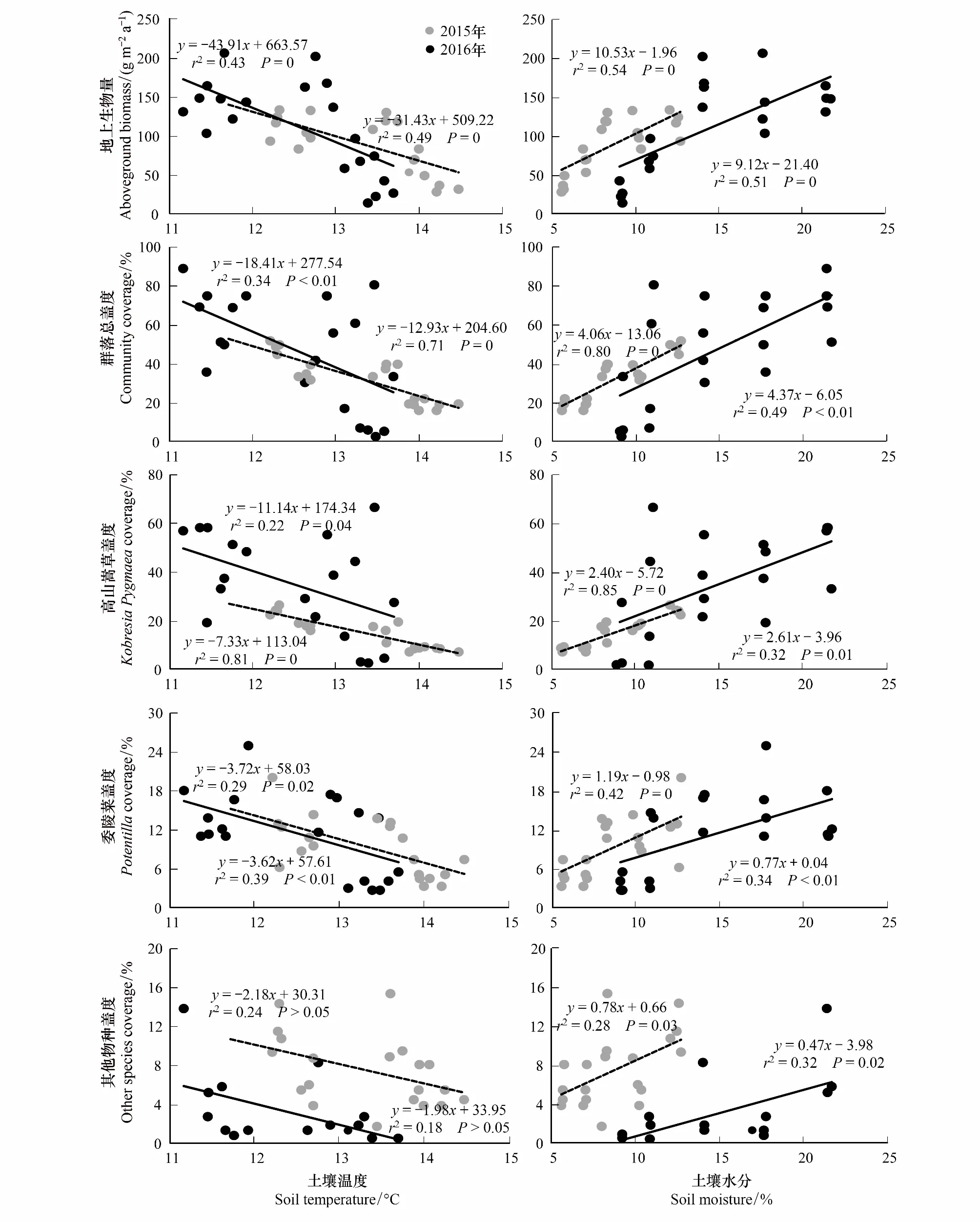
图5 线性回归分析地上生物量、群落总盖度及其组分与土壤温度和土壤水分之间的关系Fig.5 Correlations between soil temperature, soil moisture and aboveground biomass, Community coverage and composites
3 讨论
3.1 增温对地上生物量的影响
青藏高原气候严酷, 陆地表层生态系统脆弱[3],其高寒生态系统生产力及稳定性对气候变化的响应极其敏感[1]。本研究中,增温W1、W2、W3对高寒草甸植被地上生物量的累积无影响,而增温W4显著抑制了高寒草甸植被地上生物量累积(图3),其中土壤温度和土壤水分是影响地上生物量最主要的环境因子,且土壤温度越低、土壤水分越高,地上生物量越大(图5)。低温和土壤水分的有效性是限制高寒草甸植被生长及生态系统生产力的主要因素之一[31- 32],增温在一定程度上满足了高寒植物对温度的需求,进而促进高寒植物的生长[18],同时增温加快了土壤水分蒸发速率和植被蒸腾速率,加速了土壤水分的流失,进一步加剧了高寒植物的干旱胁迫,进而抑制了高寒植物的生长[33]。因此,在高寒草甸生态系统中,增温对高寒植物的生长消极作用与增温对高寒植物生长的促进作用的共同影响了增温对高寒草甸植被地上生物量累积的影响。
在全球变暖的背景下,有大量的研究通过遥感监测及田间试验模拟陆地生态系统地上生产力对增温响应,经过Meta-analyses发现在增温幅度为1.0—6.0℃的范围内,沿着增温梯度增温先是有效地促进了植物的生长再到抑制植物的生长[13- 14,23],本研究与Meta-analyses[13- 14,23]的结果一致,表明一定程度的升温会促进高寒草甸植物群落的生长,但温度升高超过一定幅度时,会导致高寒草甸植被生产力的下降,进而加剧高寒草甸的退化。其可能原因是适宜的温度升高促进了新陈代谢使其增强了植物的光合能力或导致了较高的分解作用增强了植物对矿物营养的吸收[34- 35],进而增加了地上生物量生产,同时,增温后土壤含水量的减少可能导致干旱胁迫的加剧抑制了新陈代谢或增温增强了植物呼吸作用[14,28],使其植物地上生物量累计的下降,尤其是过度增温后土壤水分流失过快,干旱胁迫严重,草甸生态系统通常以减少生物量作为代价,提高水分利用效率来适应环境[18,36]。
3.2 增温对群落特征的影响
在全球变化背景下,温度变化势必会影响植物竞争力、植物根系分布深度、根系呼吸、微生物群落的组成、植物生理、植物物候、植物养分的分配等,进而影响生态系统的结构和功能[5- 6,28]。本研究中增温导致植物群落部分物种的消失,且增温幅度越大物种消失的越多,这与李娜等[18]在高寒草甸生态系统的研究结果一致,其可能原因是从形态学上高寒植物已经进化出吸热的形态,而增温导致植物组织的温度超过了有些高寒植物的耐热极限,最终导致群落的演替[33],也可能是增温导致不同植物物种的竞争等级与相对优势的改变导致植物群落组成发生改变[37- 38],进而导致植物群落的演替,例如:增温增加了钉柱委陵菜和楔叶委陵菜的重要值、减少了二裂委陵菜的重要值(表3)。
在气候变暖背景下,温度的变化必将影响植物对养分和水分的吸收,从而间接影响植物的生长和生物量的积累[1,21,35]。本研究中,增温W1,W2,W3,W4的群落总盖度、高山嵩草盖度和其他物种总盖度均减少,且增温幅度越大,群落总盖度和高山嵩草盖度减少的越多;同时委陵菜属植物盖度在低度增温(W1、W2)无明显变化,而在高度增温(W3、W4)显著减少了委陵菜属植物盖度(图4)。高山嵩草属于典型的高寒草甸群落中的浅根植物[30],抗旱能力差[18],增温减少了土壤含水率,加剧了干旱胁迫,严重威胁了该物种的生长;同时高山嵩草盖度受环境因子土壤温度的影响,且土壤温度越大,高山嵩草盖度越小(图5),增温导致土壤温度的升高,可能直接影响了高山嵩草植物的活性[22,39],也可能抑制了地下有机质的分解[21],进而抑制了高山嵩草的生长。委陵菜属植物抗旱能力强[40],受干旱胁迫的影响较弱,进而导致低度增温无影响、高度增温抑制委陵菜属植物生长。除建群种(高山嵩草和委陵菜属植物)后的其他物种中几乎全是逃逸种,一般出现在适宜它们生长的土壤中,而增温加快了土壤水分蒸发速率和植被蒸腾速率,其土壤水分的减少可能导致干旱胁迫,严重影响了逃逸种的生长[41]。
在全球变暖背景下,对于任何一个植物群落来说,总有一些物种比另一些物种对温度升高的响应更为敏感[18,33]。本研究中,高山嵩草盖度和其他物种总盖度存在显著的年间差异,而委陵菜属植物盖度无明显的年间差异(表3,图4)。其可能原因是历史增温的滞后效应对不同植被功能群影响的差异导致的[42-43],对增温敏感的高山嵩草来说[30],在2015年生长季降水少加剧了历史增温的滞后效应,而2016年降水增多后可以有效缓解历史增温的滞后效应,而对增温不明感的委陵菜属植物来说[30],历史增温对其影响的滞后效应较弱;也可能是土壤水分有效性是限制青藏高原高寒草甸植被生长及生态系统生产力的主要因素之一[31],生长季降水的差异(2016年降水多于2015年)导致了生长季土壤水分有效性的差异,对于浅根的高山嵩草受土壤水分的限制最严重[30],而委陵菜属植物是深根植被[30],抗旱能力强[40],对干旱胁迫较其他物种不敏感,进而导致委陵菜属植物盖度无明显年际差异,这也说明在全球变化背景下相对于其他植被委陵菜属植物更稳定,这类植被在未来气候变暖的条件下可能具有更强的竞争力,其可能原因是增温后委陵菜属植物自身较大的叶面积,有利于光合速率的增加和分蘖能力的增强[44],进而促进委陵菜属植物的生长。
4 结论
通过以上的分析和讨论,得出以下主要结论:气候变暖对高寒草甸植物群落地上生产力的影响具有阶段性,低度升温没有影响高寒草甸植被的生长,但温度升高超过一定幅度时,会导致草地生产力下降,草地退化加剧。增温后,直接导致了OTC内土壤含水量下降、土壤温度增加,继而植物种间竞争关系被破坏,从而引起群落组成发生改变、植被群落总盖度和高山嵩草盖度减少,且大幅度的增温促进了这种趋势;而低幅度增温(W1、W2)对委陵菜属植物盖度无显著影响,高幅度增温(W3、W4)显著减少了委陵菜属植物盖度。同时高山嵩草盖度和其他物种总盖度存在显著的年际差异,而委陵菜属盖度无明显的年际变化,表明委陵菜属物种在全球变化背景下相对稳定,这类物种在未来气候变暖的条件下可能具有更强的竞争力。综上所述,青藏高原腹地分布广泛的草甸植物群落的生长和结构对实验增温的响应是敏感而迅速的,同时本研究为高寒草甸植被群落响应气候变化的研究提供基础数据,丰富了脆弱生态系统响应全球变化的研究。
猜你喜欢
成都信息工程大学学报(2022年4期)2022-11-18
青海草业(2022年2期)2022-07-23
农业工程学报(2022年5期)2022-06-22
海外文摘·文学版(2021年3期)2021-05-19
活力(2019年21期)2019-04-01
军事文摘(2018年24期)2018-12-26
今日重庆(2016年2期)2016-08-11
塔里木大学学报(2014年3期)2014-03-11
自然资源遥感(2014年2期)2014-02-27
自然资源遥感(2014年2期)2014-02-27
