
天山林区不同类型群落土壤氮素对冻融过程的动态响应
2019-02-20 10:43胡仲豪常顺利张毓涛
生态学报 2019年2期
胡仲豪,常顺利,*,张毓涛,陈 磊,李 翔
1 新疆大学资源与环境科学学院绿洲生态教育部重点实验室, 乌鲁木齐市 830046 2 新疆林科院森林生态研究所, 乌鲁木齐市 830063
大量的氮添加控制实验证实氮素对于维系北方温带森林生态系统的结构和功能具有重要作用[1-2]。森林土壤氮素转化动态受水热条件、微生物类群等因素变化的影响[3-4]。在秋冬季逐渐降温雪被覆盖、春季升温冰雪逐渐消融的过程中,北方高纬度、高海拔区域的土壤冻融现象频繁发生。土壤冻融交替导致的土壤团聚体破碎、凋落物分解、微生物死亡及类群变化可促进土壤氮素大量积累与释放,这部分土壤氮素能够被土壤微生物和植物直接利用或随融雪淋溶流失[5]。有关整合分析表明,冻融过程使得微生物氮和微生物C/N分别下降12.2%和8.5%,土壤可溶性有机碳和可溶性有机氮分别上升27.5%和37.3%[6]。可见,冻结环境和冻融过程可以影响土壤养分矿化、养分周转等氮素转化动态[7- 9],进而调控着凋落物分解、根系生长、群落的结构变化和系统功能稳定等生态过程[10-11]。学者们对北极苔原带、北方针叶林、北方落叶阔叶林、高山寒草地以及中纬度地区的农田等生态系统的土壤冻融过程及其生态效应已经有了较多关注[12-13],但对冻融过程土壤氮素组分、转化与流失等方面还存在一些未知问题[14- 16]。
以天山雪岭云杉(Piceaschrenkiana)为单优树种构成的天山森林在水源涵养、生物多样性保育等方面对干旱区的新疆具有不可替代的重要作用[17]。天山林区所处纬度较高,雪被覆盖时期长达4—6个月,在秋冬与冬春交替之际温度变化较大。由于雪被覆盖期间野外调查困难及方法局限,对于该区森林冬季覆雪期间土壤氮动态的研究对仍相对缺乏。
基于以上认识,本文以天山云杉森林冻融期表层土壤为研究对象,在林区不同的群落类型内采用原位土壤培养的方法,试图探讨以下问题:(1)季节性冻融及雪被如何影响天山林区土壤氮素的有效性;(2)不同类型群落的土壤氮素动态对冻融过程的响应是否存在差异。以便为深入了解冻融期森林生态系统土壤氮循环过程,解析冻融期土壤氮素损失和迁移途径,以期为天山森林可持续经营提供理论指导,同时为全球森林氮循环提供重要参考。
1 材料与方法
1.1 研究区概况
本文以中国森林生态系统定位研究网络天山森林生态系统定位站研究站(87°07′—87°28′E,43°14′—43°26′N)为依托,在天山北坡中段的雪岭云杉森林林区布设试验点。研究区属温带大陆性气候,年总辐射量达5.85×105J cm-2a-1,年均气温约为2—3℃,历年极端最高温为30.5℃,极端最低温为-30.2℃,年降水量400—600 mm,最大积雪深度为83 cm,林下土壤为山地灰褐色森林土[18]。研究区是以天山云杉纯林为主的温带针叶林,郁闭度介于0.4—0.8,其凋落物的分解是土壤中生成较厚的腐殖质的重要途径。在天山云杉森林林区,非优势种乔木主要有天山花楸(Sorbustianschanica)、天山桦(Betulatianschanica)、天山杨(Populustremula)等,林缘、林窗及林下的灌木主要有栒子(Cotoneastermelanocarpus)、小檗(Berberisheteropoda及B.nummulariaBge.)、蔷薇(Rosasp.)、忍冬(Lonicerahispida)、绣线菊(Spiraeahypericifolia)、锦鸡儿(Caraganaleucophloea)等,林下草本主要有羽衣草(Alchemillatianschanica)、老鹳草(Geraniumrotundifolium)、羊角芹(Aegopodiumpodagraria)等[19]。
1.2 监测装置布设与样品测定
选取林地、草地、灌丛各3个10 m×10 m样地(表1),每个样地布置8根原状土壤管(高20 cm,内径10 cm,共计72个),在尽量不破坏土壤的原结构情况下,将原状土埋入土壤管中。采用U盘式温度记录仪(MicroLite5032P-RH,Fourier systems,ISR)对3种不同群落类型的5 cm土层温度进行检测(记录间隔为1 h)。冻融循环次数根据土温数据来统计,规定土温在0℃以上持续3 h至0℃以下保持3 h接0℃以上至少3 h为一次冻融循环。使用超声波测距传感器(SR50A SONIC RANGING SENSOR,Campbell Scirntific,Inc.,USA)对林地和草地雪深进行测定(测定间隔10 min)。因不存在遮蔽因素,所以灌丛与草地的雪深等同。
分别在冻结初期(EFP,early-freezing period,2016年10月15日—2016年12月15日)、深冻期(DFP,deep-freezing period,2016年12月15日—2017年3月15日)、融化初期(ETP,early-thawing period,2017年3月15日—2017年4月10日)、融化末期(LTP,late-thawing period,2017年4月10日—2017年4月25日)进行采样。每次采样时,在每个各标准样地内采集1根原状土壤管。所有样品放入装有冰袋的保温箱内低温保存,迅速带回实验室,测定植物样品及土壤样品中的氮素组分。
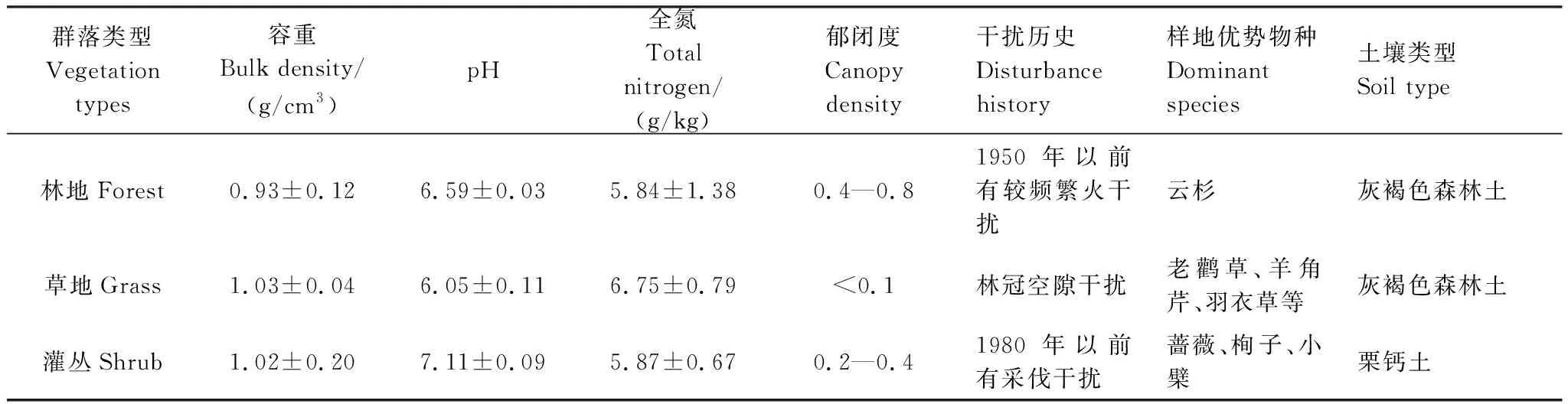
表1 样地基本信息及植物优势物种
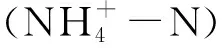
1.3 数据分析方法
土壤氮素矿化速率和植被氮素吸收速率,参考Wei[21]和katrin[22]等人的方法:
(1)
(2)
Rm=Ra+Rn
(3)
式中,Ra、Rn、Rm分别代表土壤氨化速率、硝化速率和矿化速率(mg m-2d-1)。
本文利用SPSS 17.0及SigmaPlot 13.0进行数据分析及绘图。将冻融时期和群落类型作为影响因子,通过单因素方差分析(One-Way ANOVA)和两因素方差分析(Two-Way ANOVA),判定各因子及其交互作用对土壤氮素形态及矿化速率的影响显著程度;相同冻融时期不同群落类型和相同群落类型不同冻融时期的各组分氮素含量差异采用最小显著差异(LSD)多重比较方法。
2 结果分析
2.1 土壤冻融环境动态
本研究区不同类型群落的冻融环境有一定差异,见表2。可见,冻结历时为草地>林地>灌丛,不同群落类型冻融频率变化不明显。研究区秋冬季至次年春季的地表温度见图1。草地与灌丛因林冠条件相同,因不存在遮蔽因素,使用同等雪深数据见图2。林地土壤温度在冻结初期下降迅速,深冻期土壤温度较低;草地与灌木在冻结初期土温降低缓慢冻结历时较长,而融化末期土温迅速回升,体现出雪被越厚对温度变化的阻尼作用就越明显;研究样地土壤冻结温度最小值为-5.07℃,冻融幅度及雪被波动较大(83.33—16.53 cm),主要受大气温度及林冠遮挡影响。林冠层的存在与否导致林地与灌丛草地的积雪厚度差异极大,两种条件下的积雪厚度极差可达 62.99 cm;林地覆雪时间长于草地和灌丛,林地的融雪过程较为平缓,而草地与灌丛在融雪时积雪厚度剧烈下降。

表2 不同群落类型冻融环境

图1 季节性冻融期间不同群落类型土壤日均温度动态Fig.1 Dynamics of soil temperature at the different vegetation types treatment during the freeze-thaw transitionEFP:冻结初期,early-freezing period;DFP:深冻期,deep-freezing period;ETP:融化初期,early-thawing period;LTP:融化末期, late-thawing period;GS:生长季,growth season

图2 季节性冻融期间不同群落类型土壤日均雪深动态 Fig.2 Dynamics of snow depth at the different vegetation types treatment during the freeze-thaw transition
2.2 土壤氮素各组分在冻融期的变化规律
在整个冻融时期,土壤铵态氮含量整体上呈现出与地温变化一致的规律,但不同类型群落的响应程度有差异,见图3。草地和灌丛受土壤温度影响较林地出现时空滞后,表现为先下后上升再下降的趋势。林地深冻期铵态氮含量低于灌丛和草地,各林分在融化末期的氨氮含量达到最高值(2.75—4.57 mg/kg),草地与灌丛铵态氮含量在生长季达到最低值(1.60 mg/kg,0.95 mg/kg)。相对于草地和灌丛,冻结持续时间更长(3367 h),冻融强度(-5.07℃)更大的云杉林地铵态氮含量变化更明显。
在整个冻融时期,土壤硝态氮含量对地温变化的响应比铵态氮更强烈,但不同类型群落的响应趋势较为一致,见图4。其中,从生长季进入冻融季后硝态氮含量变化最为明显的群落类型为灌丛,其在初冻期的含量为19.59 mg/kg、而在融化末期仅为6.24 mg/kg,生长季的含量为16.02 mg/kg。融化末期灌丛群落的土壤硝态氮含量较融化初期下降了64.6%。林地在深冻期硝态氮含量比其他群落类型高出43.4%—77.2%。
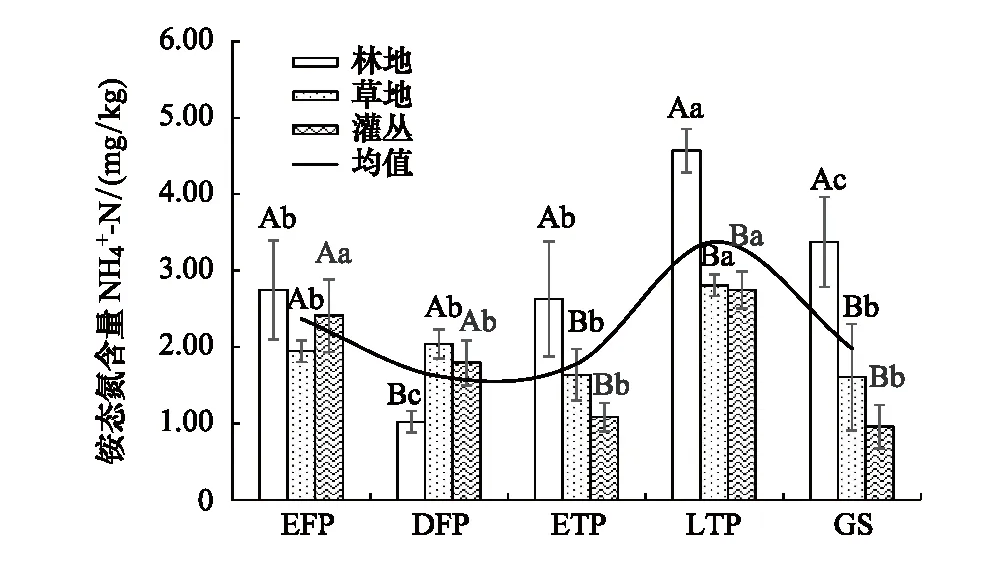
图3 季节性冻融期间不同群落类型土壤铵态氮含量动态 Fig.3 Soil ammonia nitrogen content in different vegetation types during seasonal freezing and thawing大写字母表示不同类型群落在同一时期的差异显著性(P<0.05),小写字母表示同一类型群落在不同时期的差异显著性(P<0.05)

图4 季节性冻融期间不同群落类型土壤硝态氮含量动态 Fig.4 Dynamic change of soil nitrate nitrogen in different vegetation types during seasonal freezing and thawing

图5 季节性冻融期间不同群落类型土壤微生物量氮含量动态 Fig.5 Dynamics of soil microbial biomass N content in different vegetation types during seasonal freezing and thawing
在整个冻融时期,不同群落类型的土壤微生物氮含量与地温变化规律一致,见图5。不同群落类型土壤微生物氮含量随冻融时期整体呈现先降低后升高趋势。3种群落类型均在深冻期降至低值(56.4—94.8 mg/kg),从深冻期至生长季总体表现上升趋势,林地、灌丛、草地相对深冻期分别增长了1.55、1.43、1.92倍。林地微生物氮含量除了土壤温度较低的深冻期外均在不同时期均高于其他群落类型。草地微生物氮含量(266.5 mg/kg)在生长季略高于灌丛微生物氮(261.4 mg/kg)含量。


图6 不同冻融时期土壤氮素形态Fig.6 Soil nitrogen forms at different freezing and thawing stages
2.3 冻融期氮矿化速率在不同群落类型上的差异
不同群落类型土壤氮矿化的趋势大致相同,但在不同时期净氮矿化速率差异较大,见图7。深冻期至融化初期净氮矿化速率为正值(18.84—27.54 mg m d-1),融化初期至融化末期较低,为(-17.59—-43.98 mg m d-1)。其中硝化速率在深冻期至融化初期达16.43—29.13 mg m d-1,融化末期至生长季达3.08—12.73 mg m d-1两个阶段均高于氨化速率。在深冻期至融化末期,林地净氨化速率为正值,且在融化末期至生长季较草地和灌丛硝态氮流失速率分别减缓了18.10%、63.06%。冻融时期对整体氮素矿化速率影响显著,群落类型对氨化速率影响显著,冻融时期与群落类型的交互作用对氨化速率及矿化速率影响显著。
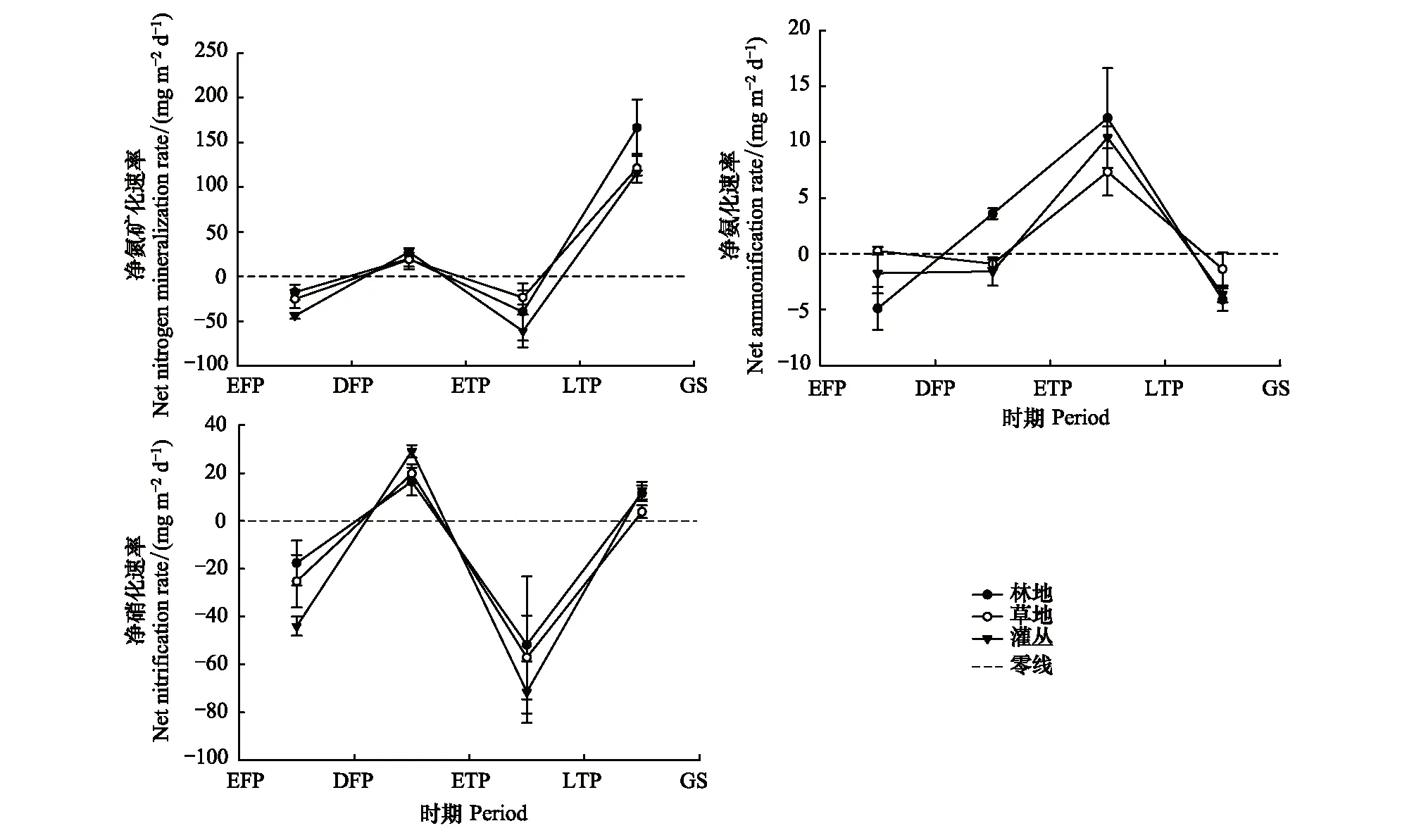
图7 季节性冻融时期不同阶段氮矿化速率Fig.7 Rate of nitrogen mineralization during different periods of seasonal freezing and thawing
3 讨论与结论
3.1 季节性冻融过程对不同类型群落土壤氮素形态的影响
以往研究表明在季节性冻融过程中土壤氮素形态主要与冻结温度,冻融幅度及冻融频率有关[23- 24],但也有近期研究发现积雪厚度在季节性冻融过程中对土壤氮素形态影响显著[25]。本研究中,冻融时期变化对土壤铵态氮含量和微生物氮含量和矿质氮含量影响显著(P<0.01)(表3),原因可能是由于冻结期的温度下降及冻融循环过程直接破坏了土壤团聚体结构,导致微生物代谢活性及酶活性的降低或致死了部分微生物[26]。此外,本工作发现土壤温度及积雪深度对氮素形态影响显著(表4),林地深冻期土壤铵态氮含量低于其他群落类型而硝态氮含量高于其他群落类型,原因可能是土壤温度的降低影响了土壤微生物群落组成进而影响到氮矿化的路径依赖,导致了硝化作用的增强与氨化作用减弱[27]。在融化初期至生长季林地氨态氮含量较高,主要因为冻融过程中的雪被移除导致了根系死亡增加了DOC含量进而影响了微生物的C源与N源,同时也与林地较高的根系生物量有关,这与Hosokawa在日本北海道西部的雪被移除及根系添加实验相吻合[28]。
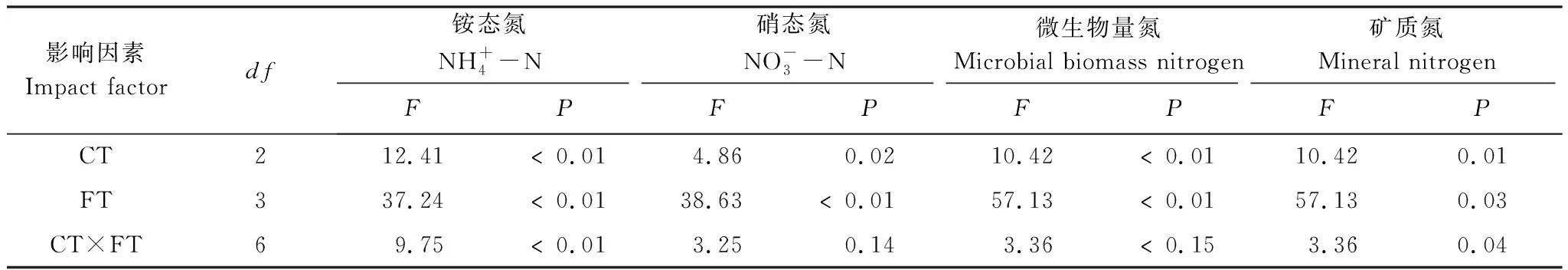
表3 冻融时期及群落类型对氮素形态的影响
CT: 群落类型,Community type; FT: 冻融时期,Freeze-thaw stage

表4 温度与积雪深度对土壤氮素形态的双因素方差分析
T:温度,Temperature; SD:积雪深度,Snow Depth
本研究中,不同群落类型均在冻结初期和融化初期释放大量有效氮素,并随冻融格局变化而出现显著差异。土壤矿质氮含量在冻结初期显著高于深冻期。可能的机制有:植被凋落物在冻结初期被大量的归还到土壤表面[29],植被凋落物的可溶性氮素释放以及冻融循环对土壤团聚体和细根的破坏作用造成了冻结初期土壤可溶性氮的大量积累[30-31]。同时,这些土壤氮素又对低温耐受性高的细菌如古氨化细菌等低温嗜冷微生物增加了有效基质[32-33],促进了土壤微生物生长和有机质矿化。另一方面,由于受雪被覆盖影响,本研究区土壤冻融速率较低,而细胞的活性很大程度取决于土壤的冻结速率。快速冻结和对消融对微生物影响较小,主要由于胞内成冰或液晶状;但细胞在缓慢冻结和融化过程中收到的损伤较大,因冰晶形成于胞外且冰晶颗粒大,对细胞器破坏力更强[34]。若冻结速率慢,则细胞可充分脱水,细胞内外渗透压平衡被破坏,此时微生物存活率与渗透压相关。相反,如果冻结速率很快,细胞内水分可以在跨膜转移前迅速冻结,则避免了渗透压对细胞的扰动[35]。这也解释了为何本研究区在冻结频率不高的情况下深冻期氮矿化量依然较低。
冻融时期对硝态氮与矿质氮影响显著(P<0.05),对铵态氮与微生物量氮影响极显著(P<0.01),因硝态氮氮含量占矿质氮含量比重较大,冻融时期对矿质氮影响也未达到极显著水平。同时,冻融时期与群落类型的交互作用对硝态氮、微生物量氮和矿质氮含量影响显著(P<0.05),对铵态氮含量影响极显著(P<0.01),这可能是气候、土壤和植被组成等多种因素综合作用的结果。
3.2 季节性冻融过程对不同类型群落土壤氮矿化率的影响
为满足自身的氮营养需求,植物在长期适应过程中会以多种方式吸收“氮源”,其中最主要的便是以主动吸收的方式从环境介质中获取矿质氮源,既铵态氮和硝态氮[36]。净氮矿化速率表征了土壤中植物可利用氮素的情况,前人研究表明土壤氨化速率和矿化速率与土壤温度成正比[37]。然而,近期研究表明,冻结或冻融循环的破坏作用可效促进冬季土壤有机氮矿化分解,并可在融化期间释放大量有效氮素,增加土壤氮矿化速率[38]。本研究中不同群落类型在深冻期至融化初期净氮矿化速率均为正值,在融化末期至融化末期温度升高矿化速率却出现负值,且硝化速率在融化初期至融化末期降低显著,见表5。究其可能的原因有:1)在高雪被覆盖的深冻期,土壤微生物在较为缓和的冻结温度下逐渐适应了微环境[39],土壤微生物活性逐渐升高,加之冻结初期土壤累计的大量有效养分被充分利用,导致矿化速率显著回升。2)天山林区融雪中后期的积雪液态含水率对土壤水热状况影响显著[40],土壤未冻水含量波动较大,而冻融季节森林土壤氮素淋溶主要形式为铵态氮和硝态氮[41],同时硝化作用与氨化作用互相影响,故而导致整体矿化速率下降。

表5 冻融时期及群落类型对土壤氮矿化速率的影响
CT: 群落类型Community type; FT: 冻融时期 Freeze-thaw stage
另外,随冻融时期变化不同群落类型对氨化速率影响极显著(P<0.01),在初冻期至深冻期原生云杉林地土壤氨化速率显著低于草地土壤与灌丛土壤,主要是由于林冠造成的雪被差异大,土壤温度低导致,而后在深冻期至融化末期氨化速率上升是由于原生林植被结构相对稳定,土壤微生物群落结构更为复杂[42],对温度响应更加敏感,加之林地雪深值较低,温度变化更为明显,故随温度上升氨化速率快速提升。
3.3 结论
以上可知,在季节性冻融期,天山北坡林区的土壤矿质氮的主要存在形式是硝态氮,土壤氮素矿化过程以硝化为主。全球气候变化情景下,温度上升不仅通过缩短冻结时间提高土壤净氮矿化速率,而且可能对高雪被覆盖下的冻融微环境造成影响,但这些现象的形成机制还需要进一步研究。由于硝态氮素在冰雪融化过程中更易流失,意味着天山林区在控制水土流失、保持土壤氮素水平、净化地表水水质等方面需要加强关注。
猜你喜欢
快乐作文(1.2年级)(2022年5期)2022-05-31
中国粮油学报(2018年12期)2018-03-19
中国公路(2017年10期)2017-07-21
中国计划生育学杂志(2017年3期)2017-06-01
中国房地产业(2016年24期)2016-02-16
百科探秘·航空航天(2015年5期)2015-12-01
健康女性(2014年8期)2015-05-12
植物营养与肥料学报(2012年1期)2012-10-26
植物营养与肥料学报(2010年3期)2010-11-16
植物营养与肥料学报(2010年4期)2010-11-06
