
lncRNA NEAT1在结核分枝杆菌感染RAW264.7巨噬细胞中的表达及作用的初步探讨
2019-03-05 11:41黄舒颖黄自坤
安徽医科大学学报 2019年2期
黄舒颖,张 诚,黄自坤,罗 清,卿 城
结核病是严重危害人类健康的全球性公共卫生问题。世界卫生组织公布的报告表明,目前全球每年约有150~200万人死于结核病,高居单一病原致患者死亡的感染性疾病之首。我国是结核病高负担国家之一,结核病患病人数居世界第三位[1]。近年来由于耐药菌株的不断增多、人口流动性加大以及HIV的蔓延,使得结核病的防治更加迫在眉睫。结核分枝杆菌(Mycobacteriumtuberculosis, MTB)是一类胞内寄生菌,主要寄生于感染受体的巨噬细胞内,因此机体抗结核免疫主要依赖于细胞免疫。遗憾的是,对MTB感染后相关免疫反应及免疫逃逸的机制至今尚未阐明,使得结核病的防治至今未再次获得突破性进展。
长链非编码RNA (long-noncoding RNA, lncRNA)是一类广泛存在的长度大于200 nt非编码RNA,缺乏开放读码框,而是利用与RNA、DNA或者蛋白的相互作用,对靶分子进行核酸降解、干扰RNA翻译等方式在转录和转录后水平对细胞的增殖、凋亡、损伤、自噬和分化等生物学过程进行调控,在免疫、肿瘤、心血管[2]等多种疾病中发挥重要作用。lncRNA NEAT1是近期研究发现的一个重要的细胞功能调控因子,在卵巢癌[3]、前列腺癌[4]等肿瘤疾病的形成、分化及转移方面发挥重要作用。重要的是,NEAT1参与了机体固有免疫过程,是体内重要的免疫正向调节因子。在感染性疾病中,NEAT1参与了人体HIV[5]、汉坦病毒[6]、寨卡病毒[7]感染过程。在免疫性疾病系统性红斑狼疮患者单核细胞中发现NEAT1表达上调,进一步研究显示NEAT1可调节THP-1细胞炎症因子白介素6(interleukin-6,IL-6)、CXCL10表达[8]。由于细胞因子IL-6和CXCL10与MTB感染后巨噬细胞炎症反应密切相关,因此,NEAT1可能参与了巨噬细胞抗MTB的免疫过程。该研究采用H37Rv菌株构建感染巨噬细胞模型,检测NEAT1在巨噬细胞中的表达,另一方面利用RNAi 干扰沉默巨噬细胞NEAT1表达,检测H37Rv感染后IL-6表达及其对H37Rv杀菌功能影响,初步探讨NEAT1在结核病免疫反应过程中的作用。
1 材料与方法
1.1材料与主要试剂RAW264.7巨噬细胞购自中国科学院上海细胞研究所;MTB标准菌株H37Rv(ATCC 27294)由本实验室保存;DMEM培养基购自美国Hyclone公司;胎牛血清购自杭州四季青公司;TRIzol 试剂和LipofectmineTM2000转染试剂均购自美国Invitrogen公司; PCR引物购自上海生工生物工程公司;IL-6 ELISA试剂盒购自上海明睿生物技术有限公司;逆转录试剂盒与荧光定量PCR试剂盒购自大连宝生物TaKaRa公司;siRNA 干扰试剂购自广州锐博公司;荧光定量PCR采用美国ABI公司(Applied Biosystems 7600系统)。其它所用试剂均为分析纯。
1.2感染模型的建立选取对数期生长的RAW264.7巨噬细胞加入24孔板,置于 37 ℃、5% CO2孵箱中培养至细胞贴壁良好后,将H37Rv按一系列感染复数(multiplicity of infection, MOI)对巨噬细胞进行感染,4 h后将胞外未感染的细菌用PBS洗去,然后继续在培养箱中培养(37 ℃、5%CO2),12 h后TRIzol提取细胞总RNA;或采用MOI=5对巨噬细胞进行感染,并于感染后4 h将胞外未感染的细菌用PBS洗去,然后置于培养箱中培养(37 ℃、5% CO2),在感染后的不同时间点(12、24、48 h)TRIzol分别提取细胞总RNA,RT-qPCR检测细胞NEAT1表达情况,ELISA技术检测培养上清液IL-6含量。
1.3总RNA提取TRIzlo法提取总RNA,具体参考试剂说明书。
1.4RT-qPCRcDNA逆转录采用TaKaRa公司逆转录试剂盒,所有操作条件按说明书进行。内参GAPDH上游引物:5′-GCACCGTCAAGGCTGAGAAC-3′,下游引物:5′-TGGTGAAGACGCCAGTGGA-3′。NEAT1 上游引物:5′-CTTCCTCCCTTTAACTTATCC-ATTCAC-3′, 下游引物 :5′ -CTCTTCCTCCACCATTACCAACAATAC-3′[8]; qPCR 反应采用 SYBR Premix EX TaqTMⅡ,使用ABI 7500 系统进行上机,具体的检测步骤及反应条件设置都按照试剂盒说明书进行。所有样品做 3 复孔,RNA相对表达量以 2-ΔΔCt方法计算所得。
1.5IL-6蛋白测定ELISA检测培养上清液IL-6浓度,具体步骤参考试剂盒说明书。
1.6siRNA细胞转染制备NEAT1 siRNA以及control siRNA(siNC),具体由上海Gene Pharma公司完成。siRNA引物序列[8]:正义序列:5′-GUGAGA AGUUGCUUAGAAACUUUCC -3′,反义序列:5′-GGA AAGUUUCUAAGCAACUUCUCACUU-3′。巨噬细胞分为3组:空白对照组、siNC转染组以及siNEAT1转染组。巨噬细胞贴壁良好后,siNEAT1或siNC转染根据LipofectamineTM2000转染试剂说明书进行操作,转染后细胞放在孵箱中培养4~6 h后更换完全培养基,转染后24 h收集细胞,TRIzol提取细胞总RNA,RT-qPCR检测siRNA敲减效果。
1.7siNEAT1对巨噬细胞IL-6分泌影响NEAT1沉默后的RAW264.7巨噬细胞,将H37Rv按MOI=5的比例对细胞进行感染,48 h后ELISA法检测细胞培养上清液中IL-6的含量。
1.8siNEAT1对巨噬细胞对H37Rv杀菌功能检测巨噬细胞分为3组:空白对照组、siNC转染组以及siNEAT1转染组;NEAT1沉默后的RAW264.7巨噬细胞,将H37Rv按MOI=5的比例对细胞进行感染,4 h后用PBS洗去胞外未感染的H37Rv,该时间点设为H37Rv感染后0 h。分别于H37Rv感染后0 h、72 h收集并裂解各组细胞,将裂解液进行系列梯度稀释,接种于含有 OADC的7H10固体培养基上,继续培养 3~4周后进行菌落计数。

2 结果
2.1H37Rv感染RAW264.7巨噬细胞后NEAT1表达和IL-6分泌情况如图1所示,MTB感染巨噬细胞后,RT-qPCR检测显示细胞内NEAT1表达显著上调,与空白对照组表达为1.0相比,12、24、48 h分别上调(5.83±1.21)、(5.25±0.98)、(4.62±0.93)倍,差异有统计学意义(F=33.55,P<0.01)。 ELISA检测表明,未感染时细胞培养上清液中IL-6的浓度为(186.43±28.09 )pg/ml,MTB感染48 h后IL-6浓度为(404.29±43.53)pg/ml,差异有统计学意义(t=11.12,P<0.01)。
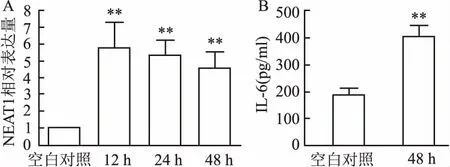
图1 MTB感染巨噬细胞后NEAT1表达和IL-6分泌情况A:NEAT1;B:IL-6;与空白对照组比较:**P<0.01
2.2NEAT1沉默后巨噬细胞IL-6分泌的变化RNAi转染成功地沉默了巨噬细胞NEAT1表达(t=16.19,P<0.05),见图2A。将沉默NEAT1的RAW264.7巨噬细胞感染MTB后收集培养上清液,采用ELISA法检测IL-6分泌水平。从图2B可见,正常对照细胞仅少量分泌IL-6 [(142.25±36.43)pg/ml],感染MTB后分泌量升高[(405.26±23.80)pg/ml],差异有统计学意义(t=12.08,P<0.01)。沉默NEAT1后的巨噬细胞,与转染对照序列细胞(siNC)比较, IL-6分泌量显著减少(t=4.45,P<0.01)。
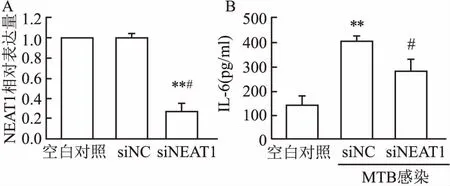
图2 ELISA检测NEAT1沉默后感染MTB的RAW264.7巨噬细胞IL-6分泌量变化
A:NEAT1;B:IL-6;与空白对照组比较:**P<0.01;与siNC组比较:#P<0.05
2.3NEAT1沉默后巨噬细胞对H37Rv杀菌能力的影响以空白敲除组作为对照组,敲低NEAT1的RAW264.7细胞为实验组,分别感染H37Rv,0.1%SDS裂解细胞并接种于7H10固体培基,最后检测荷菌量。结果显示,H37Rv感染后0 h时,siNEAT1组与对照组细胞荷菌量差异无统计学意义(t=1.03,P>0.05),提示沉默NEAT1对巨噬细胞吞噬MTB功能无明显影响。以0 h时间点胞内MTB量为基准,在MTB感染后72 h时,与对照组比较,siNEAT1处理组巨噬细胞中MTB的存活率显著上升(t=8.98,P<0.01),见图3,提示沉默NEAT1表达可显著抑制RAW264.7巨噬细胞清除胞内MTB的能力。
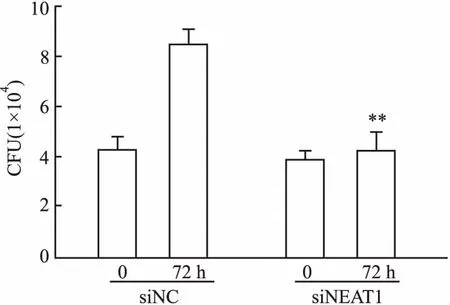
图3 siNEAT1对RAW264.7细胞杀伤H37Rv影响的检测结果与siNC 72 h组比较:**P<0.01
3 讨论
lncRNA已被证实广泛参与多种人类疾病的调控。lncRNA THRIL[9]参与巨噬细胞内TNF-α基因转录和蛋白分泌;lncRNA Cox2[10]可调控巨噬细胞内NF-κB与Toll样受体信号通路,影响炎症因子干扰素、趋化因子的表达等。此外,lncRNA在结核病免疫反应过程中的作用也逐渐引起了关注。例如lncRNA CD244可从表观遗传水平调控结核感染过程中CD8+ T细胞IFN-γ和TNF-α分泌[11]。lncRNA MEG3敲除的巨噬细胞,其BCG清除能力显著提高[12]。
NEAT1又称为MENε /β或 VINC,是从人类第11号染色体上一个被称为多发性内分泌瘤病(MEN)Ⅰ型的基因位点,由 RNA聚合酶Ⅱ转录生成,具有高度的人鼠同源性[13]。NEAT1有NEAT1-1(约3 700 bp)和NEAT1-2(约2 300 bp)两个亚型。已有的研究[8]显示,NEAT1-1是巨噬细胞的主要表达成分。因此本研究中直接选取检测巨噬细胞中 NEAT1-1表达进行相关研究。
NEAT1参与了机体的固有免疫反应调控,是一个重要的促炎分子。Zhang et al[14]发现NEAT1在 HIV-1 感染患者的 T细胞中表达上升,敲除NEAT1 则会促进 HIV-1 mRNA从细胞核到细胞质的出核与转运过程,进而促进HIV-1病毒的复制。汉坦病毒感染人脐静脉内皮细胞也会引起NEAT1表达升高,转染上调NEAT1可活化RIG-I/IRF7信号通路,促进炎症因子IFN分泌[6]。Zhang et al[8]研究发现脂多糖(LPS)刺激可提高巨噬细胞内NEAT1的表达水平,高表达的NEAT1可通过调控JNK/ERK MAPK信号通路影响巨噬细胞内炎症因子的表达。然而,目前尚未见有关NEAT1在结核病中的研究报道。
由于MTB是胞内寄生菌,在其感染机体后主要寄生于巨噬细胞内,因此抗结核免疫研究最终还是要落实到巨噬细胞上来[15]。为了明确NEAT1在MTB感染中的作用,本研究首先采用RT-qPCR技术检测了RAW264.7巨噬细胞在感染H37Rv后细胞NEAT1的表达。结果表明,MTB感染会诱导显著上调NEAT1在巨噬细胞中表达,同时IL-6分泌增加。这可能是由于免疫细胞感染MTB后,首先会迅速激活巨噬细胞,并促进包括IL-6在内的多种促炎因子释放,增强机体对MTB的杀伤能力,阻止其广泛播散。最近研究[11]表明NEAT1可通过调控JNK/ERK MAPK炎症信号通路影响巨噬细胞内炎症因子的表达。因此,在实验过程中检测NEAT1沉默后巨噬细胞IL-6分泌量变化。结果显示,siRNA敲除NEAT1后,使 IL-6分泌量下调。进一步通过胞内细菌计数实验显示,敲除NEAT1后可显著减弱RAW264.7巨噬细胞对MTB的胞内清除能力,进一步证实了NEAT1参加MTB感染后的免疫过程。
综上所述,本研究显示MTB H37Rv感染RAW264.7巨噬细胞后细胞内NEAT1表达显著升高,炎症因子IL-6分泌增加;沉默NEAT1会抑制巨噬细胞对MTB的清除能力。本研究提示NEAT1在结核病免疫反应过程中发挥了重要作用,为结核病免疫治疗提供了新的线索。
猜你喜欢
中日友好医院学报(2022年4期)2022-10-15
保健医苑(2022年5期)2022-06-10
医学概论(2022年4期)2022-04-24
现代仪器与医疗(2021年1期)2021-06-09
看世界·学术下半月(2020年7期)2020-09-10
广东农业科学(2017年10期)2018-01-25
中国资源综合利用(2017年4期)2018-01-22
中国医学创新(2017年22期)2017-11-15
现代检验医学杂志(2016年1期)2016-11-12
中国卫生(2015年1期)2015-11-16
