
饲粮添加陈化玉米和复合抗氧化剂对仔猪生长性能、抗氧化功能及肝脏损伤的影响
2019-03-11 05:23李锦英陈代文毛湘冰黄志清罗玉衡罗钧秋
四川农业大学学报 2019年1期
李锦英,陈代文,郑 萍,虞 洁,毛湘冰,何 军,黄志清,罗玉衡,罗钧秋,余 冰
(动物抗病营养教育部重点实验室,四川农业大学动物营养研究所,成都611130)
近年来我国玉米产量大幅度上升,产量的百分之七十左右用于饲料生产,剩余部分主要归入仓库临储,且伴随产量的增加,临储总量也随之上涨。我国粮食储备具有规模大和年限长两大特点,其轮换和代储的平均年限为3年[1]。如何合理高效利用储存期长的轮换玉米非常重要。前人研究表明,伴随玉米储存时间的延长,其理化性质会发生不同程度的劣化。研究发现,陈化玉米相较于正常玉米在水分、粗蛋白质和灰分[1]等养分含量上变化不显著,但储存时间长的玉米,其α-淀粉酶[2]、过氧化物酶(POD)[3]和过氧化氢酶(CAT)[4]的酶活显著下降,且胚芽部富含的多不饱和脂肪酸易发生氧化酸败[5],产生过量的脂质过氧化产物丙二醛[6]。我国GB/T20570-2015中规定玉米宜存脂肪酸值(FAV)≤65 mg/100 g,重度不宜存FAV值>78 mg/100g[7]。因脂质的氧化与水解,导致陈化玉米的FAV值上升[8],且单个挥发性物质如乙酸、己醛等及总挥发性物质含量增加[9]。储存条件良好的陈化玉米其霉菌毒素含量一般不超过国家和欧盟标准[10],但储存不当也会导致霉菌毒素的显著增加。有研究表明,畜禽饲粮中使用陈化玉米会影响生产性能。2016—2017年上半年,我国部分饲料企业因在饲料配方中采用陈化玉米而引发区域性黄膘肉现象,且猪的采食量下降、生产性能降低[11]。刘比一等发现肉鸡采食储存期长的玉米,血清抗氧化功能显著下降[12],可能破坏肝脏结构,引发炎症反应[13]。张亚茹等发现陈化玉米饲粮中添加维生素E、茶多酚等抗氧化剂,可不同程度地改善肉鸭的抗氧化能力[14]。目前,关于维生素C、维生素E、酵母硒等的抗氧化作用已进行了大量研究,其抗氧化作用已经很明确,且复合使用时具有协同效果。目前,关于陈化玉米在畜禽上使用效果的研究主要集中在家禽上,就已发表的文献看,尚无有关陈化玉米在仔猪上的研究报道,其对仔猪生长性能和抗氧化能力的影响尚不清楚。同时,在含陈化玉米的仔猪饲粮中添加抗氧化剂能否缓解陈化玉米的可能负面效应也不清楚。因此,本研究旨在用储藏4年的陈化玉米不同比例替代饲粮中的正常玉米,初步探究陈化玉米对仔猪生长性能、抗氧化能力及肝脏结构和功能的可能影响以及复合抗氧化剂的添加效应,研究结果可为养殖生产中合理使用陈化玉米作为畜禽饲料原料积累试验资料。
1 材料和方法
1.1 试验时间和地点
动物饲养试验于2017年2—3月在四川农业大学动物营养研究所科研基地仔猪代谢舍进行。
1.2 试验材料
本试验中普通玉米储存时间为1年,购自四川旺达饲料有限公司,颜色金黄,颗粒饱满且完整粒高;陈化玉米储存时间为4年,购自东北周边粮库,色泽暗淡,颗粒较饱满且完整粒低。其储存条件遵从GB/T 7415—2008标准。
1.3 试验设计与试验动物
选择24头体重相近为(7.24±0.02)kg的健康DLY断奶仔猪,按体重相近和公母各半原则随机分为4个处理(每个处理6个重复,每个重复1头仔猪),分别饲喂基础饲粮(玉米为正常)、陈化玉米50饲粮(陈化玉米50%替代基础饲粮玉米)、陈化玉米100饲粮(陈化玉米100%替代基础饲粮玉米)、复合抗氧剂饲粮(陈化玉米100饲粮+0.2%复合抗氧化剂)。试验期28 d。试猪全程单笼饲养。
1.4 试验饲粮
基础饲粮由玉米、豆粕、膨化大豆、鱼粉等组成,参照NRC(2012)7~11 kg和11~25 kg仔猪营养需要进行配制,基础饲粮组成及营养水平见表1。据试验设计用陈化玉米替代基础饲粮中相应比例的正常玉米即构成试验饲粮。复合抗氧化剂以等量替代基础饲粮中的球磨糠。
复合抗氧化剂组成:参考李彦等[15]和高亿清等[16]的研究结果,制定复合抗氧化剂组成如下:0.3 mg/kg酵母硒(纯度>95%)、100 mg/kg维生素E(纯度>95%)、200 mg/kg维生素C(纯度>95%)。
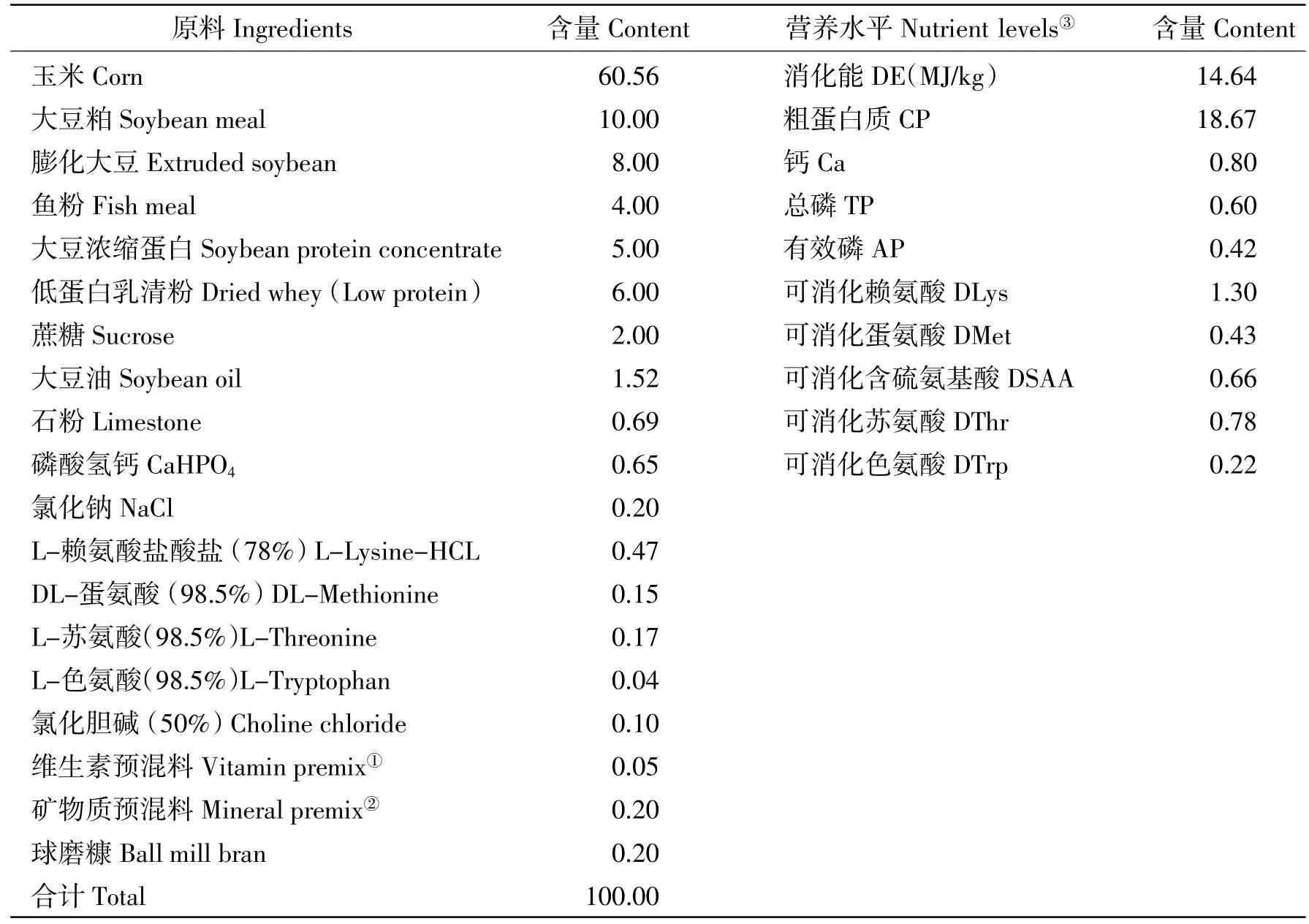
表1 基础饲粮组成及营养水平(风干基础)Table 1 Composition and nutrient levels of basal diet(air-dry basis)%
1.5 饲养管理
试验期间仔猪日饲喂3次(8:00,14:00,20:00),每次喂料采取湿拌料,以手捏成团但不出水为宜,量以吃饱后饲槽内略有余料为度,自由饮水。室温保持在26℃左右。每天打扫圈舍卫生。定期带猪消毒。
1.6 样品采集与处理
1.6.1 饲料样
将陈化玉米和正常玉米原料及各处理组全价配合饲料严格按四分法原则取200~300 g置于已知重量的瓷盘中,65℃烘干达恒重后粉碎,过40目筛,-20℃保存待测。
1.6.2 血清样
所有试验仔猪试验第28天晚20:00禁食12 h后,于试验第29天早8:00进行前腔静脉采血30 mL,室温下倾斜静置30 min后3,500 r/min离心10 min,分离血清,分装后-20℃保存待测。
1.6.3 肝脏样
试验第29天早上采血后,所有试验猪麻醉后迅速打开胸腔,取肝脏样,用预冷生理盐水冲洗血渍,滤纸吸干多余水分后分装,液氮速冻后-80℃保存待测。
1.7 考察指标与方法
1.7.1 玉米和饲粮生化特性指标
玉米和饲粮中POD和CAT活性及MDA含量采用分光光度比色法测定。所有试剂盒均购自南京建成生物工程研究所,详细操作方法参见试剂盒说明书。FAV按照SC/T3501—1996方法检测;常规概略养分含量按照张丽英[17]方法测定。
1.7.2 生长性能
试验期间准确记录每个重复每日采食量,并于试验的第1天和第29天早上对仔猪进行空腹称重,计算试猪的ADG、ADFI和F/G。
1.7.3 血清转氨酶及氧化还原相关指标
血清GPT和GOT活性,T-AOC,CAT、T-SOD和GSH-Px活性,MDA含量采用分光光度比色法测定。所有试剂盒均购自南京建成生物工程研究所,详细操作方法参见试剂盒说明书。
1.7.4 抗氧化指标测定
肝脏中CAT、T-SOD和GSH-Px活性,MDA、还原型谷胱甘肽(GSH)和PCO含量的测定采用分光光度比色法测定,所有试剂盒均购自南京建成生物工程研究所,详细操作方法参见试剂盒说明书。DNA氧化产物8-OHdG采用ELISA法检测,试剂盒购自北京冬歌博业生物科技有限公司。
1.7.5 肝脏炎性和凋亡相关基因mRNA表达量
采用实时荧光定量逆转录-聚合酶链反应(RTPCR)技术测定肝脏炎性和凋亡相关基因相对表达量。组织中总RNA的提取和质量检测参照Chen Y.等[18]的方法进行。cDNA的合成按照RT Reagent kit的说明书进行。待测基因引物序列由大连生物科技有限公司合成,引物序列见表2。用实时定量PCR仪(ABI-7900)进行测定,RT-PCR反应体系为10μL:SYBR Premix ExTaqTMⅡ(TaKaRa,Japan)5μL,上下游引物各0.4μL,cDNA 1μL,ddH2O 3.2μL。PCR扩增条件为:模板预变性,95℃,30 s;扩增,95℃,10 s;适宜的退火温度(表2),25 s,共39个循环;溶解曲线,55~95℃,温度以0.5℃/s速率提升。各目的基因的相对表达量以β-actin作为内参基因进行计算,计算方法参考T.D.Schmittgen等[19]。每个样品设定3个平行,取平均值用于数据统计分析。
1.8 数据处理与统计分析
所有数据均采用SAS8.1统计软件进行统计分析。采用一般线性模型(general linear model,GLM)对CTL组、AG50组和AG100组相关指标进行单因素方差分析,差异显著时采用Duncan法进行多重比较;对AG100组和AG100-AO组相关指标进行t检验。所有结果以“平均数±标准误”表示,以P<0.05为差异显著。
2 结果与分析
2.1 玉米和饲粮的生化特性
由表3可知,陈化玉米与正常玉米的水分、粗灰分、粗脂肪及粗蛋白质等概略养分含量无显著差异。但是,陈化玉米的CAT和POD酶活分别比正常玉米降低40.33%和47.24%,FAV和MDA含量分别为正常玉米的1.94倍和20.41倍。

表2 基因引物序列Table 2 Primer sequences of target and reference genes

表3 普通正常玉米和陈化玉米生化特性Table 3 The biochemical property of normal corn and aging corn
由表4可知,随着饲粮中陈化玉米替代比例的增加,与基础饲粮相比,含陈化玉米的饲粮其CAT和POD活性降低,而FAV增加。
2.2 陈化玉米和复合添加剂对仔猪生长性能的影响
由表5可知,与CTL组相比,AG50组和AG100组均显著降低仔猪ADFI(P<0.05),ADG分别降低8.78%和7.37%,F/G分别降低4.83%和4.12%,但差异均不显著(P>0.05)。与AG100组相比,添加复合抗氧剂能一定程度缓解陈化玉米导致的ADFI和ADG的下降,但差异不显著(P>0.05)。

表4 饲粮抗氧化酶活和脂肪酸价Table 4 The activities of dietary antioxidant enzymes and fatty acid value

表5 陈化玉米和复合抗氧化剂对仔猪生长性能的影响Table 5 Effects of aging corn and complex antioxidant supplementation on the growth performance of piglets
2.3 陈化玉米和复合添加剂对仔猪抗氧化功能的影响
由表6可知,与CTL组相比AG50组和AG100组显著降低仔猪血清GSH-Px活性(P<0.01),且AG100组显著降低T-AOC水平(P<0.01);添加陈化玉米不同程度降低血清CAT和T-SOD活性,但差异不显著(P>0.05)。与陈化玉米100组相比,复合抗氧化剂显著提高血清CAT、T-AOC活性和肝脏GSH含量(P<0.05)。
2.4 陈化玉米和复合添加剂对仔猪肝脏氧化损伤的影响
由表7可知,与CTL组相比,AG50组和AG100组显著提高肝脏8-OHdG含量(P<0.05)。与AG100组相比,复合抗氧剂显著降低肝脏MDA、8-OHdG和PCO含量(P<0.05)。
添加陈化玉米不同程度提高GPT活性,差异不显著,但AG50组和AG100组GOT/GPT值分别降低21.82%(P>0.05)和46.67%(P<0.05)。与AG100组相比,复合抗氧化剂使GOT活性增加21.95%(P>0.05),GPT活性降低28.90%(P<0.05),GOT/GPT值升高125%(P<0.01)。
2.5 陈化玉米和复合添加剂对仔猪肝脏细胞因子和凋亡相关分子表达的影响
由表8可知,添加陈化玉米和复合抗氧化剂均对仔猪肝脏促炎性相关因子TNF-α、IL-1β、NFκB、IL-6和抗炎因子IL-10 mRNA丰度,及促凋亡因子Bax和抗凋亡因子Bcl2mRNA丰度均无显著影响。
3 讨论
3.1 陈化玉米的理化特性
研究表明,随着玉米储存时间的延长,相较于正常玉米,其水分、粗蛋白质和灰分等常规养分含量的变化不显著,但α-淀粉酶、CAT及POD的酶活显著下降,且玉米胚芽部富含的多不饱和脂肪酸易发生氧化酸败,产生过量的脂质过氧化产物MDA[1-6]。脂质的氧化与水解也将导致陈化玉米的FAV上升[8]。本试验研究结果与前人相似,陈化玉米的概略养分含量变化不大,但CAT与POD的酶活大幅降低,而FAV大幅升高,呕吐毒素含量相对较高;且各处理组饲粮呈现类似变化规律。前人研究表明,当多种霉菌毒素同时存在时会协同霉菌毒素的负面效应,因而也可能会影响仔猪的采食量。因此,上述结果暗示,陈化玉米呈现脂质过氧化现象及多种霉菌毒素同时污染(虽未超标),将其用作饲料原料时可能会表现负面效应。

表6 陈化玉米及复合抗氧化剂对仔猪血清和肝脏抗氧化指标的影响Table 6 Effects of aging corn and complex antioxidant supplementation on serum and liver anti-oxidation index of piglets

表7 陈化玉米及复合抗氧化剂对仔猪肝脏损伤指标的影响Table 7 Effects of aging corn and complex antioxidant supplementation on hepatic oxidative injury in piglets
表8 陈化玉米及复合抗氧化剂对仔猪肝脏炎性与凋亡因子mRNA表达水平的影响Table 8 Effects of aging corn and complex antioxidant supplementation on mRNA abundance of inflammatory and apoptotic factors in liver of piglets
3.2 陈化玉米和复合抗氧化剂对仔猪生长性能的影响
2016—2017年上半年,我国部分饲料企业因在饲料配方中采用陈化玉米而引发区域性黄膘肉现象,且猪的采食量下降、生产性能降低[11]。Yin D.F.[20]等饲喂肉鸡陈化玉米饲粮,发现ADG和F/G显著降低。崔小燕等[21]研究表明与0年和1年的玉米相比,饲喂储存4年的玉米显著降低了1~21日龄肉鸡F/G。本试验发现,陈化玉米替代组显著降低仔猪采食量,对ADG和F/G无显著影响。原因可能有如下两个方面:第一,动物模型不同对陈化玉米的反应敏感性不同,本试验以断奶仔猪为动物模型,而此前报道中的动物模型多为家禽;第二,ADG降低不显著可能与试验时间短,样本量小有关。张亚茹等[14]在储存3年玉米中添加茶多酚和维生素E作为强效的抗氧化剂能提高采食陈化玉米肉鸭的生长性能。本试验未发现陈化玉米组添加复合抗氧化剂对仔猪ADFI、ADG和F/G的影响,原因可能与试验期相对较短,陈化玉米脂肪过氧化等特性尚未显著引发仔猪的病理变化,导致复合抗化剂在生长性能方面无显著作用。
3.3 陈化玉米和复合抗氧化剂对仔猪血清抗氧化指标的影响
脂质过氧化产生的自由基(ROS)可引起细胞膜脂质过氧化反应[22],产生的终产物MDA可与蛋白质、DNA等交联,使蛋白质活性发生改变,并损伤DNA,诱导基因突变等[23]。ROS可与生物体内的一些天然的自由基清除剂或抗氧化剂反应而终止连锁反应[24]。生物体内的抗氧化系统包括酶促防御系和非酶促反应,两者能有效清除胞外源和内源的氧自由基,维护机体抗氧化功能。刘比一等[12]发现储存期长的玉米会损伤肉鸡血清抗氧化功能。鞠雪[25]等研究草鱼摄食氧化鱼油饲料导致血清及组织抗氧化酶活下降。本试验结果与前人相似,表明陈化玉米饲粮可能导致机体抗氧化能力下降。王显慧[26]等在肉鸡饲粮中添加维生素C、维生素E均显著提高血清抗氧化酶活性。张亚茹等在陈化玉米饲粮中添加维生素E、茶多酚等抗氧化剂,发现抗氧化剂可不同程度地改善机体抗氧化能力[13]。本试验添加复合抗氧化剂可能改善陈化玉米组仔猪机体的抗氧化功能。
3.4 陈化玉米和复合抗氧化剂对仔猪肝脏的影响
肝脏具有解毒、免疫、凝血及胆汁生成与排泄等功能,其结构功能的正常对猪的生长健康起着重要作用。王永庆[27]发现饲料中添加氧化油脂可能对团头鲂肝脏组织结构、脂质代谢和蛋白质合成功能造成损伤。A.Ammouche[28]等饲喂氧化葵花籽油可以改变大鼠肝脏脂肪酸的组成,甚至会出现仅存在于氧化油脂中的反式脂肪酸。A.Kode[29]研究表明,加热氧化的葵花油会导致大鼠肝脏磷脂脂肪酸组成发生剧烈变化,细胞膜流动性变强。孙得发[11]等发现生长育肥猪采食陈化玉米饲粮后,肝脏发生肿大现象。崔小燕[10]饲喂储存期长的玉米显著降低了鸡肝脏GSH-Px活性和GSH含量。本试验未发现陈化玉米饲粮组对仔猪肝脏抗氧化酶活的影响,原因可能与试验期较短有关。复合抗氧化剂的添加可显著提高仔猪肝脏GSH含量,表明复合抗氧化剂能改善仔猪肝脏抗氧化功能。陈科全等[30]研究表明饲料中MDA可显著升高血清GPT的含量。本试验结果与前人相似,表明陈化玉米可能通过升高血清GPT对肝脏功能造成影响。饲粮MDA含量不仅会引起机体脂质过氧化损伤,同时又可与蛋白质和DNA交联、互作[31-32],造成机体蛋白质和DNA氧化损伤。本试验中AG50组和AG100组显著提高肝脏8-OHdG水平,添加复合抗氧化剂显著提高仔猪肝脏GSH含量,显著降低肝脏其他氧化损伤产物含量。以上结果表明,陈化玉米可降低仔猪机体抗氧化能力,且造成不同程度的氧化损伤,且相对于直接在饲粮中直接添加氧化鱼油或MDA而言,陈化玉米中所含MDA的量相对低,且以玉米为外在载体,对机体的应激效果可能不及直接添加该物质,且病理现象的发生是一个长期沉积的过程。前人报道育肥猪饲料中采用陈化玉米引起黄膘肉现象,这可能就是一个长期存积的效果。本试验短期饲喂虽并未发现其对仔猪生长性能及抗氧化能力和肝功能等造成显著损伤,但是已经显著降低仔猪采食量,导致机体抗氧化功能的下降和肝脏的氧化损伤,这对仔猪后期的生长发育与健康实际已经带来隐患;添加复合抗氧化剂能一定程度上改善仔猪机体抗氧化能力并缓解机体受到的氧化损伤。该结果为陈化玉米用作畜禽饲料原料的合理使用积累了试验依据。
此外,研究发现陈化玉米可能会破坏肉鸡肝脏的组织结构并引起炎性反应[13]。在病理状态下,自由基是致炎介质,能激活NF-κB和调节炎症相关因子[33]。本试验发现陈化玉米添加组不显著影响肝脏炎性因和凋亡因子mRNA的表达丰度;复合抗氧化剂的添加也无显著作用。表明短期内陈化玉米对仔猪肝脏的炎性损伤和细胞凋亡无显著影响。
4 结论
本试验条件下,与正常玉米相比,陈化玉米中CAT、POD酶活下降,FAV、MDA含量上升。饲粮添加陈化玉米显著降低仔猪ADFI,对ADG和F/G无显著影响,导致机体抗氧化功能的下降和肝脏的氧化损伤。添加0.2%复合抗氧化剂一定程度改善机体抗氧化能力,缓解陈化玉米导致的肝脏氧化损伤。
猜你喜欢
动物营养学报(2022年9期)2022-10-12
动物营养学报(2022年8期)2022-09-01
包装工程(2022年5期)2022-03-21
中国饲料(2021年17期)2021-11-02
兽医导刊(2020年11期)2020-12-28
中国茶叶加工(2020年3期)2020-10-21
中国粮油学报(2019年4期)2019-07-12
中国药理学与毒理学杂志(2015年3期)2015-12-16
时代英语·高三(2014年5期)2014-08-26
