
应用定向进化获取受体高亲和性hEGF突变株E40V及其特性的评价*
2019-03-29 10:52孙宇杰杨南燕欧锦新李飞黄雪莹王庆环刘健王倩倩刘昕张擎
中山大学学报(自然科学版)(中英文) 2019年2期
孙宇杰,杨南燕,欧锦新,李飞,黄雪莹, 王庆环,刘健,王倩倩,刘昕,张擎,3
(1. 中山大学生命科学学院生物工程中心,广东 广州 510275; 2. 深圳市美沃西联生物科技公司,广东 深圳 518133; 3. 中山大学深圳研究院,广东 深圳 518057)
人表皮生长因子(human epidermal growth factor,hEGF)是一种由53个氨基酸构成的单体多肽。该蛋白作为有效的促有丝分裂因子,可以和细胞表面受体EGFR特异性结合并在整个表皮和上皮组织中起作用,包括表皮,肺上皮,成纤维细胞,神经系统,角质层等[1-3]。EGF与EGFR的复合物晶体结构中显示EGF与EGFR-EC通过3个结合区域发生相互作用[4]。该相互作用可以刺激EGFR胞内发生磷酸化,从而诱发下游信号通路,导致细胞发生增殖、分化、迁移等。hEGF基因的失调与某些癌症的生长和发育有关[5]。EGFR及其突变体可作为癌症治疗的靶标[6-7]。之前有研究表明具有增强促有丝分裂能力的EGFR激动剂可用作伤口愈合剂[8-9],EGF蛋白本身对于表皮损伤和溃疡也具有良好的修复功能。所有这些发现都表明EGF-EGFR是作为受体介导的药物分配系统的重要靶标,对其进行研究是相当重要的。因此需要对EGF进行改良以获得更优越的EGFR激动剂。EGF在室温条件下能稳定7 d左右,但其生物活性在第2周和第3周分别降低35%和40%(p< 0.01)[10]。水溶液中EGF稳定性更差,并且该降低幅度同EGF稀释的程度呈正相关[11]。EGF在不同温度下存在着不同的失活机制,但总体来说EGF具有在酸性环境下更易降解,随温度升高或浓度降低降解率增加的特点。EGF的降解主要包括羧基端氨基酸的水解,Asp11的琥珀酰亚胺化,Met21的氧化以及Asn1的脱氨基等,不同的条件(温度,pH,荧光等)下会以某些反应占主导地位[12]。相对于稳定性来说,靶向性主要针对于临床应用,针对于正常组织的EGF在促进正常上皮细胞增殖的同时也有可能促使癌细胞的发展[13-14]。以上两个问题限制了EGF的实际应用。
近年来,许多新方法被广泛应用于蛋白质和多肽的优化,如基于序列-结构功能域的统计分析[15],分子定向进化等。统计分析属于理性设计,需要大量的统计计算并且要求充分了解蛋白质的基序和功能之间的对应关系。而DNA定向进化技术是一种通过定向压力筛选目的基因,使目的基因在较短时间内得到富集并丰富其多样性,最终得到所需DNA以及蛋白的方法[16-17]。其中,Coco等[18]通过简并引物的方法获得了一个人-鼠来源全交换DNA重组文库,并从中发现了一个具有94倍亲和力和123倍促增殖活性的突变子EGF21。然而,该突变株对于酸性环境较低的耐受能力以及靶向性较低限制了其具体的应用。
因此,为了获得具有高亲和力,高促增殖活性,高稳定性的EGF突变子,我们采用交错延伸技术(staggered extension process,StEP)[19]和噬菌体展示技术[20]对人源和猪原EGF进行定向筛选,获得了hEGF突变株E40V。其与EGFR结合的ELISA结果以及EGFR竞争抑制实验的数据的表明,其值为103.1±19,最大结合数Bmax为1.302±0.086。在促增殖活性实验中,E40V对于NIH3T3细胞的促增殖能力强于野生型,而对于A431细胞则与野生型相仿。为了解突变E40V对hEGF稳定性的影响,我们通过圆二色谱检测了hEGF和E40V在不同pH条件下二级结构的变化。其结果表明,E40V对于酸性的耐受性略逊于野生型,碱性耐受能力与野生型相仿。本研究不仅为短肽的定向进化提供了一个实例,也为了解EGF结构与功能之间联系打开新的视角,同时为EGF的实际应用打下基础。
1 材料和方法
1.1 材料和试剂
文章所涉及酶均购自TaKaRa,所涉及抗体均购自Abcam公司。噬菌体M13购自American Research Products。全长hEGF,pEGF cDNA克隆自cDNA文库,其序列参考自GenBank(NM_001963.3 and NM_214020.1)。大肠杆菌E.coliDH5α和BL21 (DE),质粒pET32a,pCANTAB5E购自Clontech;质粒pMD18T购自TaKaRa。细胞株A431和NIH 3T3购自中国科学院生物化学与细胞生物学研究所。胎牛血清(FBS),培养基DMEM,胰酶,EDTA均购自Gibco公司。其余试剂如无特殊说明均购自SIGMA。
1.2 StEP重组文库的构建
通过BLAST分析显示人,猴,猪,大鼠和小鼠之间的序列同源性分别为98%,85%,68%和76%(Error! Reference source not found.-A)。只有一个氨基酸差异,人和猴EGF的DNA序列之间没有足够的多态性。因此,猪EGF的DNA序列是上述五种哺乳动物中唯一满足家族重组至少80%同源性要求的哺乳动物。因此,重组文库由人EGF(hEGF)和猪EGF(pEGF)基因构建。
该实验步骤按照Zhao等[21]描述的步骤进行。将hEGF和pEGF基因作为模板,用以下4条引物进行PCR:primer 1,5′-TAGGCCCAGCAGGCCAAT AGTGACTCTGAATGTCC-3′; primer 2, 5′-CCGGCGGCCGCGCGCAGTTCCCA CCACTTCAG-3′; primer 3, 5′-TAGGCCCAGCAGGCCAATAGTACTCTGAAT GCCCGC-3′; primer 4, 5′-CCGGCGGCCGCGCGCAGCTCCCACCATTTCAAG-3′。其中,primer 1 和3 含有SfiⅠ位点(下划线部分),primer 2 和4 含有NotⅠ位点(下划线部分)。之后用DNA交错延伸的方法来生成基因重组。PCR的第一次循环分别包含primer 1,primer 3或者 primer 2,primer 4。之后反应以如下条件进行60个循环:94 ℃, 1 min;45 ℃,10 s;72 ℃,3 s。其PCR产物用作第二轮PCR的模板,此时所用引物为: primer 1,primer 2,primer 3,primer 4。使用如下条件进行基因全长的扩增:94 ℃,5 min;94 ℃,30 s;50 ℃,45 s;72 ℃,1 min,0 cycles;2 ℃,7 min。
将回收产物交叉组合后等比例混合配制PCR反应体系模板,进行StEP第二轮扩增。之后将PCR产物回收后连入pMD19T载体并转化至E.coliDH5α。之后从筛选培养基中随机挑取24株阳性克隆,使用简并引物进行菌落PCR,对扩增产物进行SSCP凝胶鉴定后[22],得到重组文库。
1.3 噬菌体展示文库的构建及筛选
hEGF及其突变株的噬菌体展示文库是按照Abdolalizadeh等[23]的描述构建的。将重组文库连接至噬菌体展示载体pCANTAB5E后将其转化至E.coliTG1中。使用辅助噬菌体M13KO7对展示文库进行噬菌体拯救后得到噬菌体展示文库。
筛选步骤参考Abdolalizadeh等[23]的描述进行。步骤大致为将A431细胞与噬菌体展示文库封闭孵育后,使用PBS缓冲液和洗脱缓冲液(含w=0.1% Tween 20的PBS)充分冲洗,之后侵染E.coliTG1。将侵染后的TG1进行噬菌体拯救后,完成1轮筛选。
1.4 EGF突变株文库受体亲和力测定
在筛选进行了4次后,将所得文库稀释103倍并平铺在SOBAG平板,使最终滴度约为2×106pfu。之后从SOBAG平板上随机对20个菌落进行噬菌体拯救,以拥有pCAN-hEGF和pCANTAB5E质粒的噬菌体为对照组,通过ELISA测定20个菌落与A431细胞株的亲和力,其步骤按Zhao等[24]的描述进行,最后用酶标仪(Labsystems Dragon Wellscan MK3)在490 nm处检测文库亲和水平。
1.5 EGF高亲和力突变株的表达纯化
含有C端His tag标签的野生型EGF及其突变株在E.coliBL21中表达和纯化的过程是按照之前描述的进行的[25]。所有质粒均进行了测序验证。本实验采用BCA法[26]测定纯化后蛋白浓度。最后通过Tris-tricine PAGE胶和Western Blot观察表达情况。
1.6 突变株对受体亲和力测定
本研究通过使用ELISA来研究蛋白对受体的亲和力水平。首先通过ELISA绘制出突变株和受体结合的饱和曲线,其步骤参考Shoko等[27]的描述进行。其中,一抗选用生物素标记的抗his-tag抗体(1∶3 000),之后使用亲和素标记的辣根过氧化物酶(1∶5 000)与一抗特异性结合。显色时每孔加入0.5 μL 4.0% OPD, 5 μLφ= 30% H2O2,1.0 μL 10×酶作用底物buffer,8.5 μL ddH2O,室温孵育20 min,最终使用2 mol/L H2SO4终止反应,并用酶标仪(Labsystems Dragon Wellscan MK3)检测492 nm下的吸光值。实验重复3次。使用GraphPad Prism 7.0 for Windows[28]对数值进行Specificbinding拟合,得出EGF突变株-EGFR饱和曲线。之后进行了EGF突变株竞争性抑制EGF结合EGFR的实验,将EGF突变株和EGF梯度稀释后分别互相混合作为结合酶,之后步骤与ELISA实验相同,实验重复3次。
1.7 EGF及其突变株的促增殖,促迁移活性检测
1.7.1 MTT法检测促增殖活性 MTT 分析过程按照Hardwicke等[29]的描述进行。其步骤大致为将NIH3T3细胞株饥饿处理4 h,把EGF及其突变株按照浓度梯度溶于w=0.2% FBS DMEM中,将其添加至NIH3T3细胞中,孵育24 h。之后按照MTT细胞增殖和细胞毒性检测试剂盒(Sigma-Aldrich LLC)说明进行。使用酶标仪(Labsystems Dragon Wellscan MK3)在490 nm处检测结果活细胞数目。实验设置3组重复,用GraphPad prism 7.0软件分析数据。
1.7.2 肿瘤细胞集落形成实验 将约200个A431细胞接种于Φ 35培养皿中,将EGF及其突变株按10 nmol/L浓度分别溶于w=1% FBS DMEM中并与细胞混合,另设一组空白对照,不加任何药物处理。每隔3 d取样并用结晶紫染色法对细胞染色,之后拍照并统计集落形成数目。
1.7.3 EGF及其突变株的促迁移活性检测 将NIH3T3细胞,A431细胞以5×105个/皿的密度接种在Φ 35培养皿中,之后对平板均匀划线并弃去培养基,使用预冷PBS缓冲液洗去划下的细胞后分别加入EGF及其突变株的w=2% FBS DMEM溶液(10 nmol/L)。37 ℃,φ=5% CO2培养48 h后拍照并统计结果。
1.8 圆二色谱法测定不同pH下EGF及其突变株的稳定性
圆二色谱实验参照Kang等[30]所描述步骤进行。用圆二色谱上样缓冲液(不同pH)将hEGF,突变株分别稀释至10-6mol/L,室温孵育15 min。将1 mL缓冲液加入比色皿中确定基线后,于比色皿中加入1 mL样品,使用分光偏振计JASCO J-720(Tokyo,Japan)读取圆二色谱值,其扫描速度为50 nm/min,波长间隔0.2 nm,模式为连续扫描,实验重复3次。pH更换时重新加入相应pH缓冲液再次确定基线。
之后使用上样缓冲液(pH8.0)稀释样品至10-6mol/L,之后操作步骤同上,在测定时设定变温范围为25-85 ℃,每10 ℃读数一次,每次读数前,预温1 min使样品温度稳定。圆二色谱数据使用CDtoolX软件[31]进行分析。
2 结果与讨论
2.1 重组文库的构建以及特性测定
使用w=3%琼脂糖凝胶和w=15%SSCP凝胶检测易错PCR和StEP产物的结果如图1-A所示。将StEP产物导入pMD19T质粒后将其转化至E.coliDH5α并进行文库容量计算。其容量为7×104pfu。之后在12个随机克隆进行菌落PCR后,使用SSCP对文库突变率进行测试,其结果如图1-B,表明初始文库发生了重组和位点突变,总突变率为0.21%,与之前报道的结果相近[32-34]。
2.2 噬菌体展示文库的筛选
由于EGF突变株被展示在M13噬菌体衣壳蛋白表面,且具有相对独立的空间构象和活性,因此表达相应EGF突变株的噬菌体可结合至EGFR过表达的A431细胞株表面,而EGF突变株对于受体的亲和力便可用A431细胞株表面结合噬菌体的量来表示。同时,初始的EGF重组噬菌体展示文库由于文本量过大,因此需要使用A431细胞株对EGF突变株进行筛选,来富集EGFR高亲和性突变株。
通过4轮的淘洗,拯救,文库得到了一定的富集,最终滴度为3×108pfu/mL。之后使用抗M13噬菌体衣壳蛋白的单克隆抗体进行ELISA来量化EGF突变株的亲和力。其结果如图1-C,可以看出,筛选后文库对于EGFR大多具有良好的亲和力,其中有4株的亲和力略高于野生型EGF,有6株的亲和力远低于野生型hEGF,其余样本处于A值在0.233-0.263之间。以上结果表明EGFR亲和株得到较为有效的富集。
2.3 EGF突变株的表达
对上述4个样品进行测序,结果表明4组均发生了不同程度的亲本交换。在4株样品中,存在一株突变株以hEGF为模板,第40位出现Glu突变为Val(以下均表示为E40V)。由于其位置位于整个蛋白的D-loop区域,与两个β折叠紧密相邻,且和EGFR的3个残基发生相互作用,推测该突变可能会对蛋白功能产生较大影响(见图2-A黑色方框标示位置),因此选择E40V进行后续研究。之后我们对野生型EGF及E40V在原核生物中的表达情况进行了探索,其结果表明野生型EGF和E40V均能在E.coliDH5α中正常表达,其western blot的检测结果如图2-D。
2.4 E40V对EGFR亲和水平测定
E40V对于A431细胞株表面EGFR结合的饱和曲线如图2-B 。其值为103.1±19,最大结合数为1.302±0.086。符合Yu等[35]的报道。
而在竞争性抑制实验中,hL8P,pP8L对hEGF的竞争抑制实验结果表明(图2-C),E40V对野生型EGF构成竞争抑制,这表明E40V与野生型结合受体相同。

图1 StEP重组文库,噬菌体展示文库,噬菌体文库对EGFR的亲和力水平Fig.1 Detection of StEP recombinant library, phage display library, phage library affinity level for EGFR(A)StEP单向文库;(B)SSCP检测文库多样性;(C)ELISA检测噬菌体展示文库的EGFR亲和性;虚横线为hEGF样品的吸光值(A) StEP unidirectional library;(B) SSCP detection of library diversity;(C) ELISA detection of EGFR affinity of phage display libraries.The dashed line is the absorbance of the hEGF sample
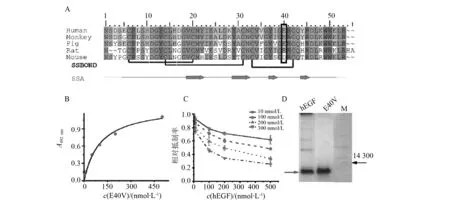
图2 hEFG及其突变株的表达及对于EGFR的结合水平Fig.2 Expression of hEFG and its mutants and their binding activities to EGFR(A)不同来源EGF的氨基酸序列比对结果。SSBOND表示二硫键,SSA表示EGF的二级结构示意图,黑色方框为EGF第40位氨基酸的保守性;(B)E40V-EGFG饱和曲线;(C)E40V在不同浓度下与hEGF的竞争结合曲线;(D)E40V和hEGF蛋白Ni-NTA纯化后western-blot结果(A) Amino acid sequence alignment results of EGF from different sources. SSBOND represents a disulfide bond in EGF structure, SSA represents the secondary structure of EGF;(B) E40V-EGFG saturation curve;(C) A competitive binding curve of E40V to hEGF at different concentrations;(D) Western-blot results of E40V and hEGFafter purification by Ni-NTA affinity chromatography
2.5 EGF突变株促细胞增殖活性检测
对EGF突变株进行了MTT检测衡量其对正常细胞有丝分裂的促进的作用,同时通过集落形成实验分析了其对肿瘤细胞集落生长的作用。
在MTT法中,我们通过将各试验组按照等浓度梯度稀释,检测其在不同浓度下对NIH3T3细胞的促增殖能力。如图3-A所示,E40V在全浓度范围对于NIH3T3细胞的促增殖能力均强于野生型,T检验分析显示其促增殖活性相对于野生型有显著优势(p> 0.05)。
之后为了验证EGF及其突变株对肿瘤细胞集落形成的影响,我们以肿瘤细胞株A431作为研究对象,进行了集落形成实验,其结果如图3-B-C。向体系加入10 nmol/L药物刺激7 d后,所有药物组均有集落形成,E40V集落大小与野生型相似(图3-C),形成数量略高于野生型,但是差异不明显(图3-C)。说明EGF促进肿瘤细胞集落形成的能力与其促增殖活性呈正相关。10 d后所有药物组的集落形成能力开始趋同,差异性减弱(结果未显示)。
2.6 EGF突变株促迁移活性检测
我们除了考察E40V的促增殖活性外,还考察了其对肿瘤细胞A431和正常细胞NIH3T3的促迁移活性的影响。结果表明(图4),在药物刺激48 h后,A431细胞均发生明显迁移,但各药物组之间差别不大。Huang等[36]的研究表明,EGF促增殖与促迁移的信号通路并不相同,即促增殖活性与促迁移活性并不是直接相关,本实验结果也从宏观层面证实了此观点。对于NIH3T3细胞,各药物组间也显示出较强的促迁移能力,而E40V相较于野生型,其促迁移能力略有增加。
2.7 圆二色谱检测EGF及其突变株的稳定性
通过圆二色谱检测了在不同pH下,hEGF和E40V的二级结构变化情况。由于EGF结构仅由β片层和loop区域构成,因此检测β片层在整体结构中的比率就能很好的反映出该蛋白在相应环境中结构的稳定情况。其结果如图5所示。在环境pH为中性和酸性情况下,E40V的稳定性远不如野生型,但是随着环境pH的逐渐升高,E40V的稳定性逐渐提升。在pH8.0下,其稳定性已与野生型持平,甚至略有提高。虽然E40V对于酸环境的耐受能力不如野生型,但是相较于于其他已报道的突变子,其耐酸能力有较大提高[18]。这可能由于所突变氨基酸为人,猪亲本发生的互换所致。
3 结 论
近年来,随着研究技术的进步,人们不再满足于简单的获取与利用EGF,分子水平上的研究和改造也已开始进行。有报道用工程学方法改造EGF取得了一定的效果,体外筛选发现了对EGFR亲和性、促生长活性增强的突变体。但这些报道大都未进一步从结构与功能或者应用表达方面有进一步的研究。而通过使用不同物种的短DNA片段进行改组已获得一些促增殖能力增强的EGF突变体[18]。虽然这种方法可以有效地构建嵌合体库,但是该过程太复杂而不能方便地操作。因此,我们的目标是通过改进传统方法,创建一种简单,方便的EGF进化方法,以获得某些特性强化的EGF突变株。
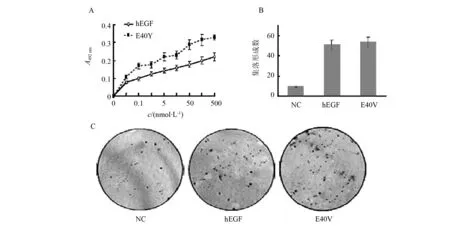
图3 hEGF及E40V对细胞增殖能力的影响Fig.3 Effect of hEGF and its mutants on cell proliferation(A)MTT检测EGF及E40V对NIH3T3细胞的促增殖能力;(B, C)hEGF及E40V对于A431细胞株集落形成的影响 (A) MTT assay for the proliferative capacity of EGF and E40V on NIH3T3 cells;(B, C)Colony formation assay of hEGF and E40V on A431 cell line
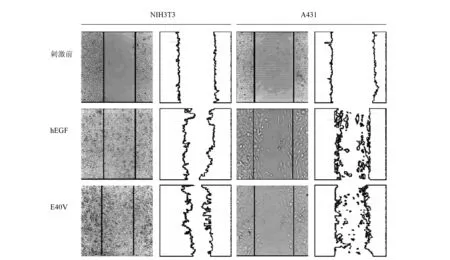
图4 A431,NIH3T3细胞分别受到hEGF及E40V刺激48 h后的迁移情况Fig.4 The ability of migration of different cells after 48 h stimulation with hEGF and its mutants
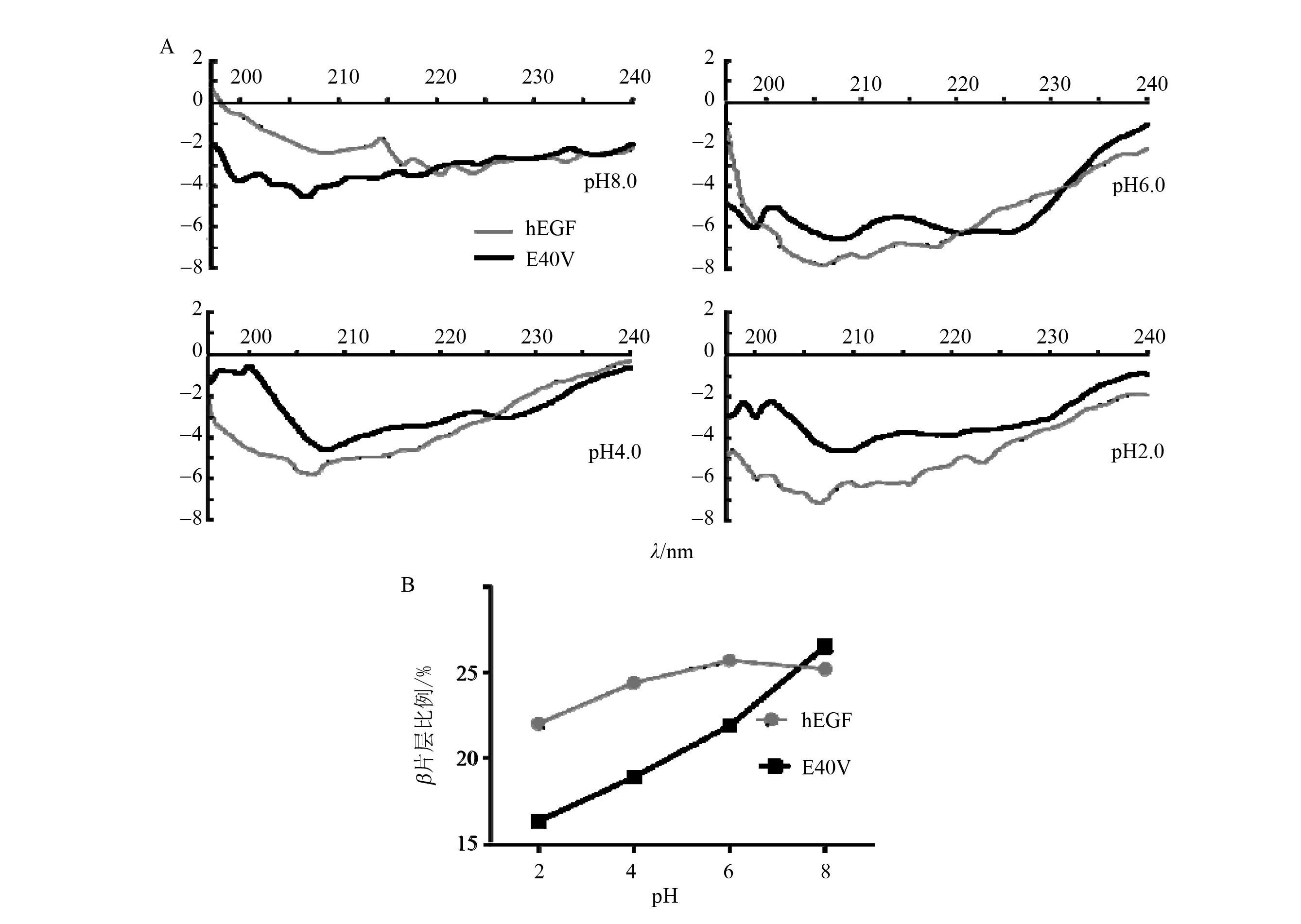
图5 EGF,E40V在不同pH下的CD谱线图以及β片层在整体结构中的比率Fig.5 CD line plot and the ratio of β-sheet in the overall structure of EGF, E40V under different pH
通过BLAST分析显示人与猴,猪,大鼠和小鼠之间的序列同源性分别为98%,85%,68%和76%。人和猴EGF DNA序列之间没有足够的多态性。因此,猪EGF的DNA序列是上述5种哺乳动物中唯一满足家族重组至少80%同源性要求的哺乳动物。因此,重组文库由人EGF(hEGF)和猪EGF(pEGF)基因构建。由于易错PCR和StEP法构建重组文库特别适用于小分子,经常用于进化蛋白质[37-39]。据报道,StEP是一种简单而有效的重组方法,因为它可以在一个步骤中实现父模板之间的交叉[19]。此外,为了增加重组,巢式引物和简并引物已用于StEP[37, 40]中。而在本研究中选择简并引物是因为EGF的DNA序列太短而不能进行巢式PCR。与最近添加适当浓度的Mn2+或高浓度Mg2+相比,通过使用Mutazyme®-mutagenesis试剂盒更容易实施易错PCR。前一种方法可以方便地在PCR过程中引起基因突变,同时也可以导致突变偏倚。然而,后者在随机突变[37, 41]中表现良好。在这项研究中,易错PCR用不同浓度的Mg2+和Mn2+进行测试(数据未显示)。发现易错PCR在1.0 mmol/L Mg2+,0.25 mmol/L Mn2+环境下具有良好的PCR产率和较高的突变率,因此我们在研究中使用相同的实验条件实施易错PCR,获得了重组率较高的重组文库。
在用噬菌体展示和ELISA筛选的研究中,最常见的筛选剂是抗体[42-44]。人们普遍认为噬菌体展示淘选活细胞可以用于筛选拮抗抗体抗肿瘤细胞[45],而且,最近一些研究人员已经开始尝试选择抗细胞表面标记糖蛋白的抗体或配体[46]。此外,还有针对活细胞的噬菌体展示平移技术[47]。由于基于A431细胞的筛选系统是研究EGF和EGFR之间相互作用的常用工具,因为A431在其细胞表面上有过量表达EGFR(每个细胞(1-2)×106个受体)[48-49]。同时,对活细胞的筛选可以模拟体内环境以获得更多功能性蛋白质。因此在本研究中,我们通过使用A431细胞淘选噬菌体展示文库,获得了功能性突变EGF,其具有高于野生型的受体亲和力。
本研究中我们利用定向进化技术将hEGF第40位Glu突变为Val。这与镰状细胞病的突变相似。在镰状细胞病中,突变Glu→Val发生在β-珠蛋白基因的第6个密码子上,改变了排列,导致血红蛋白的携氧能力下降,并进一步改变了红细胞的外观[50]。而在本研究中,EGF第40位的氨基酸在不同物种之间高度保守(图2-A),这表明其在EGF功能的发挥上具有一定的影响力。进一步的结构观察表明,第40位氨基酸于第3个二硫键附近的第3个残基环(图2-A),并且与EGFR的第15和12位氨基酸发生相互作用。从结构中可以看出,E40V相较于野生型,其侧链基团空间占位要大于野生型,而这样的突变可能更适合EGFR第12-15位氨基酸所形成的配合位点(图6)。同时,尽管40Glu突变为Val未导致蛋白质的主要结构改变,但侧链特征的差异将对EGF的亲和性和促有丝分裂活性产生影响。首先,从酸性到中性氨基酸(Glu→Val)的变化会改变局部离子环境,并且第40个氨基酸紧密接近结合关键位点(Arg41)43,44,所以改变可能影响Asp355(EGFR)和Arg41(EGF)侧链之间的盐桥和Arg41(EGF)侧链,提供范德华力接触Tyr13(EGF)和Phe357(EGFR)侧链。其次,在细胞表面pH(pH7.4)下E40V的亲和力低于hEGF野生型,在细胞质pH(pH6.0)下对EGFR具有更强的结合亲和力(图2)。这意味着当复合物内化到细胞质中时,E40V-EGFR复合物比hEGF-EGFR的复合物显示出更强的稳定性,从而导致E40V-EGFR复合物可以减少配体耗尽和受体下调。因此,复合物E40V-EGFR可以比hEGF-EGFR在细胞表面上提供更持久的刺激,以获得增加的促有丝分裂效力。此外,CD光谱的结果显示E40V随着酸度或温度的升高而易于变为线性化,尽管在pH8.0下突变体EGF和hEGF之间没有主要的结构改变。由于突变发生在C-末端附近,随机Loop在整体结构比率的增加也可归因C-末端的线性化,而这些结果有助于增强EGF45的促有丝分裂能力,需要进一步的实验来检验这些假设和组织或动物实验以测试突变体的活性。
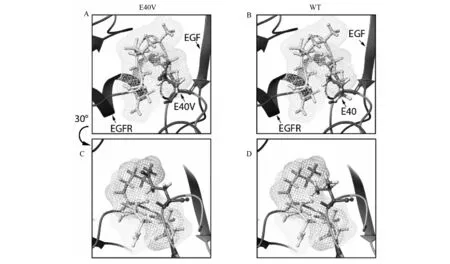
图6 E40V及hEGF的第40位氨基酸与受体的配合情况Fig.6 The binding of the 40th amino acid of E40V and hEGF to the EGFR 网状结构表示EGFR的第15和12位氨基酸的表面空间,其余氨基酸已在图中标出。C、D为将蛋白模型以y轴逆时针轴旋转30°得到
猜你喜欢
湘潮(上半月)(2022年7期)2022-12-06
昆明医科大学学报(2022年2期)2022-03-29
植物保护(2021年4期)2021-11-12
昆明医科大学学报(2021年8期)2021-08-13
昆明医科大学学报(2021年3期)2021-07-22
猪业科学(2021年3期)2021-05-21
幽默大师(2020年10期)2020-11-10
中西医结合肝病杂志(2020年2期)2020-10-27
科学24小时(2020年4期)2020-05-14
中华诗词(2019年1期)2019-11-14
