
羊草根茎克隆形态可塑性研究进展
2019-04-04 03:23白乌云侯向阳武自念任卫波赵青山
草业科学 2019年3期
白乌云,侯向阳,武自念,任卫波,赵青山,3
(1. 中国农业科学院草原研究所,内蒙古 呼和浩特 010019;2. 内蒙古师范大学化学与环境科学学院,内蒙古 呼和浩特 010022;3. 国家牧草改良中心,内蒙古 呼和浩特 010010)
生殖是生物繁衍后代延续物种最基本的行为和过程,是种群形成、发展和进化的核心问题,也是生物群落和生态系统演替的基础[1]。在强烈竞争和严酷环境条件下,与有性繁殖相比,克隆繁殖在选择上具有一定的优势,因此,在许多群落,如稀树草原、草地、极地及高山,植物主要以无性繁殖为主。中国东北样带草甸草原、典型草原和荒漠草原中,无性系植物所占比例分别为42.13%、64.11%和66.15%[2],在典型草原和荒漠草原区,克隆植物的相对重要值显著高于非克隆植物。作为一种重要的克隆植物类型,根茎型克隆植物在资源异质性生境中表现出特有的适应方式,根茎在植物无性繁殖、克隆分株间信息交流和物质交换、预测资源斑块质量等方面具有重要意义[3]。
克隆形态(clonal morphology)是指由克隆性决定的形态性状及其衍生性状[4]。当克隆植物生长在不同资源水平的生境内,其构件大小和生长结构会对环境条件产生反应,从而表现出形态可塑性(morphological plasticity)[5-6]。 对 不 同 生 境 的 适 应 是可塑性变化的重要原因。可塑性能够调节自然选择对植物的影响,为植物应对快速的环境变化提供对策,实现快速适应新环境。无性系植物的可塑性被认为是适应异质环境的重要对策之一。有关无性系植物地上构件的形态(叶、茎杆、穗等的形态,株高、节长和生物量等)可塑性方面已有广泛研究,形成了较为统一的指标和方法体系,获得了公认的结论,如长期放牧和刈割导致草地植物矮小化[7]。但无性系植物地下构件的可塑性研究起步较晚,主要有克隆构型、根茎形态、根茎芽和分株等方面的研究,目前已成为无性系植物生态学研究的热点之一。
草地植物的可塑性是其应对各种逆境的策略之一[8]。羊草(Leymus chinensis)是禾本科赖草属多年生根茎型克隆植物。羊草根茎无处不分枝,无处不生根,被称为可以通过根茎行走的游击型植物。羊草通过不断产生根茎芽及克隆分株来维持其营养繁殖,通过年复一年的螺旋扩张[9],占领新生境,成为草地群落的优势种。羊草根茎在养分储存与运输、信号传导、克隆繁殖、克隆整合、适应逆境和分工合作[10]等方面发挥着重要功能。羊草根茎克隆形态可塑性是其能够耐受干旱、盐碱和放牧等多种胁迫,在不同生境中广泛分布,大面积存在于欧亚草原的重要原因之一[11]。羊草根茎克隆形态可塑性研究中常用的形态指标如表1所列。揭示羊草独特的根茎克隆形态可塑性规律,对羊草草原保护、草原生态建设、栽培草地建植、退化草地改良和发展草食畜牧业生产等具有重要意义。为此,以根茎为研究目标,就有关盐碱胁迫、土壤、水分等环境因子和群落环境、草地利用方式等生态学因子,对羊草根茎克隆形态可塑性影响的研究结果进行归纳与总结,以期阐明羊草根茎克隆形态可塑性的生物学机理,为草地植物地下器官的研究提供理论依据。

表 1 羊草克隆繁殖研究常用根茎形态指标Table 1 Morphological index of rhizome commonly used for clonal reproduction of Leymus chinensis
1 环境因子对羊草根茎克隆形态可塑性的影响
1.1 盐碱胁迫
羊草又名碱(硷)草,是耐盐碱的优良牧草。土壤pH(碱)和含盐量(盐)对羊草克隆生长均有影响,相对于碱,盐对羊草克隆生长的影响更为复杂,但碱的破坏作用比盐更大[12]。根茎能帮助羊草应对盐碱胁迫[12-13]。围绕羊草根茎对盐碱胁迫的克隆形态可塑性已有大量野外和实验室模拟研究。
研究表明,随着土壤盐碱化程度的加重,羊草根茎平均间隔子长度和分枝角度呈增加趋势,呈游击型克隆构型,以逃离或避过盐碱生境斑块[14-19]。节间距方面,刘滨硕等[14]和李静等[15]得出了与史丽等[16]、杨智明等[17]和沈振荣等[18]相反的结论。刘滨硕等[14]发现土壤盐碱会抑制羊草根茎伸长生长。沈振荣等[18]发现在盐碱白僵土上,羊草根系呈分散型分布;而在风沙土上,趋向于直线游击型分布格局;盐碱白僵土上的分蘖株数比风沙土多。羊草根茎克隆特性适应盐碱生境潜能的发挥顺序依次为间隔子长>分枝角度>节间距[17]。根冠比[15]和比根长[19]的增加均有利于羊草适应重度盐碱化生境。
羊草有黄绿型和灰绿型两种叶色明显不同的趋异类型。羊草表型的差异不仅有广泛的环境基础,而且不同基因型的羊草性状差异及性状可塑性变异受遗传因素控制,为其应对外界环境条件变化提供保障[11]。在相同盐碱胁迫强度下,灰绿型羊草根茎的平均节间距、间隔子长度和分枝角度均显著大于黄绿型羊草[20],趋于游击的克隆构型可使灰绿型羊草分株迅速跨越或逃离恶劣生境。在内蒙古兴安盟的羊草草甸、羊草草甸草原[21]和松嫩平原沙土和盐碱土生境中[22],黄绿型羊草的营养繁殖力更强,单位面积内羊草分蘖株数和根茎长度均是黄绿型大于灰绿型。
与天然草地的复杂环境相比,盐碱胁迫模拟试验可更好地反映出盐碱单因子对羊草根茎克隆生长的影响。羊草根茎在盐胁迫条件下能够有效地吸收Na+,根茎与根一样参与Na+的吸收和运输,甚至羊草根茎中的Na+含量高于根[23],根茎吸收Na+的效率高于根[24]。单纯高pH或高盐度对羊草生长的危害远低于二者的协同效应,pH对根茎生长的抑制作用随盐度增大而加强[25]。颜宏等[26]发现当 NaCl浓度超过 600 mmol·L-1、Na2CO3浓度超过175 mmol·L-1时,羊草分蘖率分别下降30%和48%,平均根茎数分别下降58%和55%。随盐浓度的增大,芽库的储备和分株输出明显减少,根茎生物量则增大[27]。
羊草通过形态和生理可塑性应对盐碱胁迫[28]。羊草根茎除了产生分蘖茎和储存同化物外,还具有物质运输和信号传导功能。重度盐碱胁迫抑制羊草根茎生长,表现为根茎总长度变短、条数减少;但根茎生物量投入增加;间隔子长度和分枝角度增加,以迅速逃离或避开恶劣生境斑块。
1.2 土壤
土壤不仅影响羊草地上部形态学特征[29-30],也影响根茎克隆特性。除土壤盐碱化程度外,土壤养分、土壤质地和紧实度等也影响羊草根茎克隆生长。
野外调查表明,克隆植物在群落中的相对重要值与土壤全氮含量呈负相关关系[2]。在养分条件较好时,植物倾向于将生物量较多地分配给地上部分,在养分条件较差时则倾向于分配至地下部分。同质生境中,羊草分株随机分布;而在异质生境中,在营养最丰富的斑块中生长出更多的分株[31]。羊草通过缩短根茎节间距,增加不定根数来获取生境中有限的资源[15]。尽管羊草草原生产力主要受N元素限制,但在一定条件下也受P元素限制,N供应水平的变化影响羊草对P的吸收[32]。羊草总生物量和地上生物量主要受N元素限制,地下生物量主要受P添加的影响,尤其在中、高N水平下,P添加会显著降低羊草地下生物量[33]。增加养分(尤其是N元素)可促进羊草地上部生长,地下部能量分配相对减少。随N和P添加梯度的增加,羊草根冠比显著降低[34]。羊草根茎对N有吸收和转运的作用[35]。土壤全氮含量高则羊草根茎短,根茎分枝角度和子代根茎数目受影响不大[36]。施肥(NH4NO3和KH2PO4)时羊草平均分枝角度无明显变化,但平均节间距和间隔子长度缩短,即克隆构型趋于更加密集[37]。宋彦涛等[38]发现低和高N水平下羊草比根长较高,而比茎重相对稳定。N一方面抑制根茎伸长,影响羊草克隆的扩展能力;另一方面促进克隆分株的输出,增强种内不同植株对光的竞争,从而影响羊草遗传多样性、空间占领能力和生活史对策[39]。
羊草根茎主要分布在地下5-15 cm深处,长可达3 m,分枝可达4~5级,其在土壤中的横走延伸和克隆分株的输出受土壤质地和紧实度的影响。土壤紧实度与土壤质地和牲畜踩踏密切相关,其对羊草根系生长的影响可能超过盐碱胁迫[40]。在羊草种群的腊熟期,扎龙湿地林间风沙土、沙土和草甸土3种不同生境中,羊草种群分蘖株数分别为 1 080、1 290 和 1 011 株·m-2,芽数分别为 1 410、2 113 和 2 279 个·m-2,根茎累积长度分别为 7 343、8 000 和 6 948 cm·m-2,羊草种群单位面积内根茎长度和根茎生物量表现为沙土>林间风沙土>草甸土>中度碱土[41-42]。风沙土人工种植羊草有利于分蘖芽与根茎芽的发生,地上产草量大于盐碱白僵土[43]。根茎形态对土壤的可塑性主要表现在根茎长度上,随着沙质成分的增多,羊草、赖草(Leymus secalinus)、早熟禾(Poa annua)和无芒雀麦(Bromus inermis)4种根茎禾草的根茎长度相应增长,分枝强度和节间距无明显变化,因此伸长根茎的觅食行为可能主要是通过增加根茎节数来实现的[44]。王莹[31]则发现土壤中沙质成分的多少对羊草的根茎条数、根茎芽数、间隔子长度和初级根茎长度均无显著影响。
综上所述,除营养因素外,疏松的沙质土,有利于羊草根茎的伸长,羊草可通过根茎扩展固定沙土;紧实的土壤虽然形成的芽数多,但其中成长为分株的比例相对较少。
1.3 水分
当前正在发生的全球气候变化已经引起了全球水热格局的重新分配,从而将对全球植物产生重要影响。据预测,我国干旱和半干旱区在未来将变得更加干旱[45]。羊草广布于干旱和半干旱气候区,是欧亚大陆草原区东部草甸草原及干旱草原的重要建群种,开展其根茎对干旱的适应与调控机制的研究具有重要意义。
羊草根茎密度对土壤水分梯度的响应非常平缓[46],但无性系营养枝和根茎芽密度等数量特征受影响较大,与沙丘种群相比,低平原种群营养枝和根茎芽平均密度分别高出50%和30%[47]。羊草根茎直径和分枝角度与土壤含水量分别呈显著正相关和负相关关系,干旱抑制羊草节间伸长,根茎分枝角度的可塑性受水分影响较大[48],该结论与以往克隆植物分枝角度的可塑性很弱甚至不对环境变化发生显著反应的报道[49]不同,表明羊草通过增大根茎分枝角度来减少分株对水分的竞争。
降水量的增加可补偿放牧对植物的损害[50],降水量更是决定群落分布的重要因子之一。在相对干旱的克氏针茅(Stipa krylovii)群落中,羊草根茎长,根茎节数多;在相对湿润的贝加尔针茅(S.baicalensis)群落中,羊草根茎长度缩短,根茎节数减少,节间距相对稳定[51]。在松嫩草甸草原自然生境中,增雨区羊草根茎生物量显著高于对照区和减雨区 ,增雨区总子株输出数量、分蘖株数量、根茎节子株数量和总芽数均显著高于减雨区(P<0.05)[52]。内蒙古退化草地上,随着水分梯度的下降,羊草主根系的长度、体积和面积逐渐增加[53]。
实生羊草幼苗的根和根茎都能对聚乙二醇(PEG)模拟的干旱胁迫产生响应,且根茎对干旱胁迫的反应更敏感[54]。杨理等[55]和任文伟等[56]在实验室条件下得出了相似的结论,即水分促进羊草地下根茎(根茎数、根茎节数和长度增加)和无性系分株(分蘖数、根茎芽数和无性系分株数增多)的分化,缺少水分时羊草甚至不能分化出无性系分株。干旱胁迫时,羊草无性繁殖方面的性状(如分蘖数和根茎芽数等)的稳定性高于营养生长方面的性状(如株高和叶长等)[56]。
由于觅食行为,羊草增加根茎长度和分枝角度、减少分株输出,以游击型的克隆构型,减少分株对水分的竞争来应对干旱胁迫;而作为重要的环境因子,水分又可促进羊草根茎伸长和分化。调节芽分化出新枝条的几率可能是羊草对水分的适应对策之一。羊草群落生境(年降水量350 mm、湿润系数0.3、典型栗钙土地带)被认为是羊草种群无性系分化的最优生境,羊草群落中的羊草无性系最突出的根茎克隆特点是根茎长、根茎节数多、节间距大、根茎干重大[51]。
可见,羊草可通过增加间隔子长度,以迅速逃离盐碱胁迫斑块;贫瘠的土壤环境中,羊草节间距缩短,根茎节数增加,以增加须根来获取养分;疏松的土壤和充足的水分有利于羊草生长和克隆繁殖(表2)。

表 2 盐碱、土壤、水分对羊草根茎克隆形态的影响Table 2 Effect of saline-alkali, soil and water on the clonal morphology of Leymus chinensis rhizome
1.4 其他环境因子
此外,羊草根茎克隆形态对温度、光照和海拔也表现出可塑性反应(表3)。
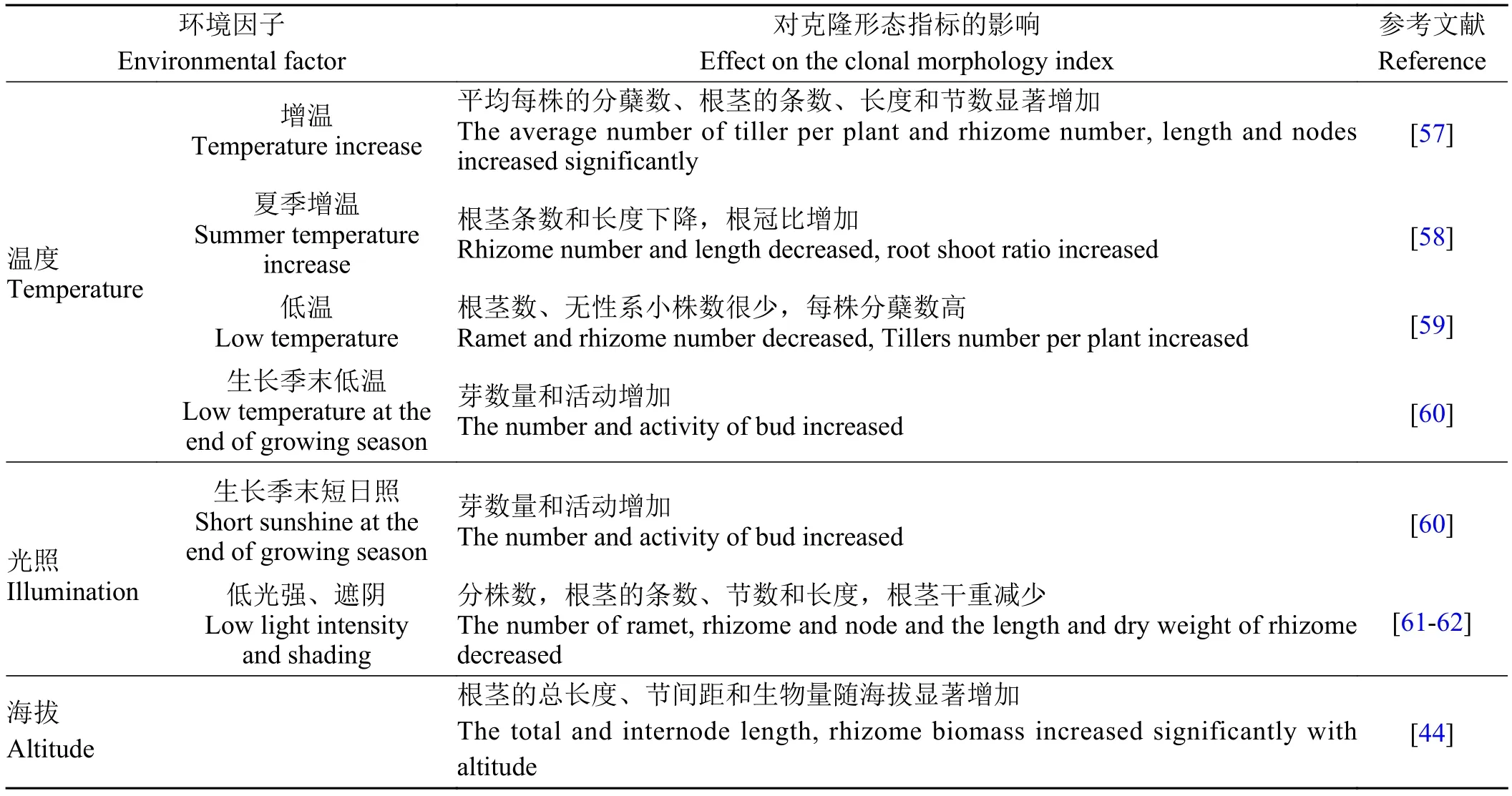
表 3 温度、光照和海拔对羊草根茎克隆形态的影响Table 3 Effect of temperature, illumination and altitude on the clonal morphology of Leymus chinensis rhizome
温度是促进羊草分蘖的主要因子之一。羊草无性系的分化与高温下的生长天数呈明显正相关关系,加温时平均每株的分蘖数、根茎的条数、长度和节数均显著增加[57]。羊草对高温有较强的适应性,甚至在高温已经抑制羊草生物量积累的情况下,仍然对无性系生长和分化有一定的促进作用。夏季增温导致羊草根茎条数和长度下降,根冠比显著增加[58]。在低温下(秋冬、冬春季)生长的羊草每株分蘖数明显高于其他时期生长的羊草,但根茎数和无性系小株很少;而高温(5月以后)下生长的羊草分化出大量的根茎和无性系小株,但分蘖的出生率小于低温处理时的出生率[59]。长时间低温显著抑制羊草根茎和无性系小株的分化,而对分蘖无明显抑制作用。羊草更新芽数量及活动与低温和短日照相关联,伴随着短日照和低温的来临,芽的数量和活动都有所增加[60]。
低光强或遮阴显著抑制羊草根茎生长和无性系小株的分化[61-62];羊草克隆分株数、根茎的条数、长度、节数和干重减少,且在春季影响更大。
大多数羊草根茎克隆形态可塑性研究是在水平空间进行的,对不同海拔下的研究较少。群落中克隆植物的相对重要值与海拔呈正相关关系[2]。在大青山呼和浩特地区羊草群落中,随海拔上升,羊草根茎总长度、节间距和生物量呈显著增加趋势;间隔子长度则对海拔无明显反应[44]。
2 生态学因子对羊草根茎克隆形态可塑性的影响
2.1 群落环境
生物群落是由水热条件主导的环境与生物相互影响、相互制约而形成的组合体,群落性质由组成群落的各种生物对环境的适应性所决定,不同群落的植物组成和生境条件不同[63]。羊草根茎克隆特性受其所在群落的植物密度、植物组成和羊草种群在群落中优势度的影响。
Schimd和Harper[64]研究发现,不同生长模式的克隆植物间的种间竞争具有密度和克隆格局依赖性:克隆构型为密集型的雏菊(Bellis perennis)在高密度环境中克隆扩展能力强,具入侵性,而在低密度环境中克隆增长受抑制;游击型的夏枯草(Prunella vulgaris)则与之恰恰相反。何念鹏等[65]发现随种群密度的增加,羊草根茎节间距和间隔子长度逐渐减小,分枝角度增大,羊草游击型特征减弱。
群落植物组成对羊草根茎克隆构型也有影响。研究发现,羊草 + 寸草苔(Carex duriuscula)群落中羊草短节间比羊草 + 虎尾草(Chloris virgata)群落和羊草 + 星星草(Puccinellia tenuiflora)群落多,可能是该群落中与羊草相伴生的寸草苔亦是一种根茎型植物,且其根茎分布范围与羊草相似,对土壤营养及土壤水分的竞争激烈所致[66]。根茎分枝角度的显著差异使得在内蒙古锡林河流域草地和沙地中羊草根茎克隆构型趋于游击型,而在林地趋于密集型[67]。羊草单优势种群落单位面积内羊草分蘖株数量和生物量分别是杨树(Populus)群落样地的近3倍和2倍,单位面积内根茎总长度和总生物量比杨树群落中的稍高[68]。羊草群落中羊草的克隆构型在果后营养期较生殖期有更游击的趋势;在羊草 + 鸡肠儿 (Herba centipedae)群落、羊草 + 寸草苔群落、羊草 + 虎尾草群落和羊草 + 星星草群落中,果后营养期的羊草克隆构型均较生殖期时趋于密集[66]。
在群落演替过程中,群落组成、盖度(或密度)、各成员型的相对优势度等不同。在水生植物群落演替初期,群落中密集型无性系植物较多;随着演替的进行,生境条件的改善,密集型无性系植物减少,而游击型无性系植物增加[69]。群落的物种多样性与生境状况存在密切关系,还与群落中的植物密度、克隆植物的重要值、植物克隆构型和克隆植物种群内部的调节机制有关:在低植物密度生境中,物种多样性随密集型和游击型克隆植物相对重要值的增加而增加;在高植物密度生境中,高度密集的密集型克隆分株阻止其他物种的定居,容易形成局部垄断格局,从而降低群落物种多样性;当游击型克隆植物进入高密度生境中时,它会借助于其分株的空间扩展优势迅速在生境中取得优势地位,排斥其他物种,导致物种多样性降低[2]。在群落水平上,长寿命的无性系植物可能“冻结”群落空间格局,减缓演替进程或产生偏途顶级。羊草草甸水淹后的恢复演替过程初期,在具有充分生长空间而没有种间竞争的风沙土上,在羊草 + 杂类草群落和羊草 + 寸草苔群落中,羊草无性系种群营养繁殖数量增长规律为幂函数,在具有种间竞争的碱化草甸土上,仍可以通过指数形式迅速增加无性系子代数量,扩展无性系生态位空间[70]。松嫩平原羊草群落随着演替的进行,单位面积内羊草分蘖株数和根茎总长度均呈上升趋势[71]。
可见,群落环境对羊草根茎克隆特性的影响是复杂的,从某单一角度去解释群落中羊草根茎克隆特性可能是片面的。深入研究密度导致的种内竞争和伴生植物导致的种间竞争对羊草根茎克隆特性的影响,对于栽培草地建植和人工辅助草地恢复具有重要指导意义。
2.2 草地利用方式
放牧和割草是草地主要的两种利用方式。草地生态学研究中不仅要研究放牧条件下群落的演替过程及方向,而且还应加强放牧条件下草原植物的生态适应对策研究[72]。羊草对长期放牧和刈割最显著的可塑性响应就是矮小化[7]。近年来,放牧和刈割对羊草根茎克隆形态可塑性影响研究日渐增多。
2.2.1 放牧
放牧逆境包括家畜的采食和践踏,是一个综合的环境梯度。放牧强度和放牧制度影响草地植物功能性状、群落结构和生态系统功能[50, 73]。植物功能性状对放牧的响应受植物功能类群的调节[74],根茎和匍匐茎游击型克隆植物比丛生密集型克隆植物更耐牧[73, 75]。草原植物的营养繁殖和生长方式及其在不同放牧强度下的适应性或对策性变化,是其能否适应放牧而维持生存的重要因素。
放牧强度影响天然羊草种群无性系繁殖的更新途径。随着放牧强度的增加,羊草营养枝密度和根茎芽密度均显著下降,特别是重度放牧后下降迅速。与轻度放牧相比,适度放牧、重度放牧和过度放牧时,羊草种群营养枝密度分别下降42.02%、74.84%和90.33%;根茎芽密度分别下降56.21%、92.26%和96.39%,极度放牧条件下几乎没有生殖枝的分化和根茎芽的形成[76]。Wang等[77]发现,中度放牧强度下分蘖株数和芽数最多。侯牡丹等[78]则认为轻度放牧最有利于植物的营养繁殖,不放牧次之。青秀玲[79]和祁永[75]发现,放牧后羊草间隔子长度显著缩短,游击性减弱,萌蘖节上的枝条数无显著差异。短花针茅(Stipa breviflora)荒漠草原群落中羊草根茎长度随放牧强度的增加而缩短;根茎节数表现为重牧>中牧>对照>轻牧;萌蘖枝条数表现为轻牧>对照>中牧>重牧[78]。宋彦涛等[80]发现中度放牧处理下,羊草根系的深度、总表面积、总投影面积、总体积和平均直径都高于轻牧和重牧处理,因此认为与丛生型草本植物相比,根茎型草本植物的粗根直径和根系体积是反映其对放牧干扰最敏感的指标。长期放牧导致分蘖数减少,节间距缩短,改变叶、根和根茎的生物量分配[77]。
放牧不但改变羊草种群赖以生存的土壤环境,使土壤盐碱化加重,更重要的是放牧时由于家畜的采食和践踏导致羊草种群无性繁殖体数量下降。过度放牧会导致羊草根茎长度、间隔子长度和节间距缩短,羊草根茎游击型变弱,耐牧性降低。过度放牧也会抑制根茎芽的形成和子株的输出[76],使羊草无法维持和延续种群正常增长,进而加快羊草草地退化速度。可见持续过度放牧对草地的损害是极大的。推迟放牧、降低放牧压力[81]和改善放牧制度和管理[50]可减轻放牧对草地的不利影响。
2.2.2 刈割
随着舍饲和集约化养殖的推进,刈割对牧草根系和分株生长的影响成为草食畜牧业研究的热点之一。
割草样地冬性枝条的存量显著高于围栏样地[71],表明割草不仅促进根茎芽的形成,而且极大地促进芽输出形成冬性枝条,但对成株与成根茎两种更新芽的形成却有较大的不良影响[82]。刈割频次对羊草根茎克隆也有影响。随刈割频次的增加,羊草间隔子长度和根冠比呈单峰曲线,而节间距则呈上升趋势,刈割处理下的羊草分枝很少[66]。西辽河平原不同利用条件下,分蘖株总数围封割草利用地段最多,封育网围栏外围羊草单优势种群落地最少;根茎总长度在围封生长后期割草利用地最长,全年放牧地最短;冬性枝条和芽在全年放牧利用地最多,围栏封育的割草地最少[83]。刈割条件下,羊草根茎的营养繁殖力大于分蘖节的营养繁殖力[71]。适度刈割可使羊草将更多物质分配给地下器官,游击特征增强[66]。在刈割草地上,羊草种群以形成大量根茎顶端芽成株的无性繁殖对策维持种群增长[82]。
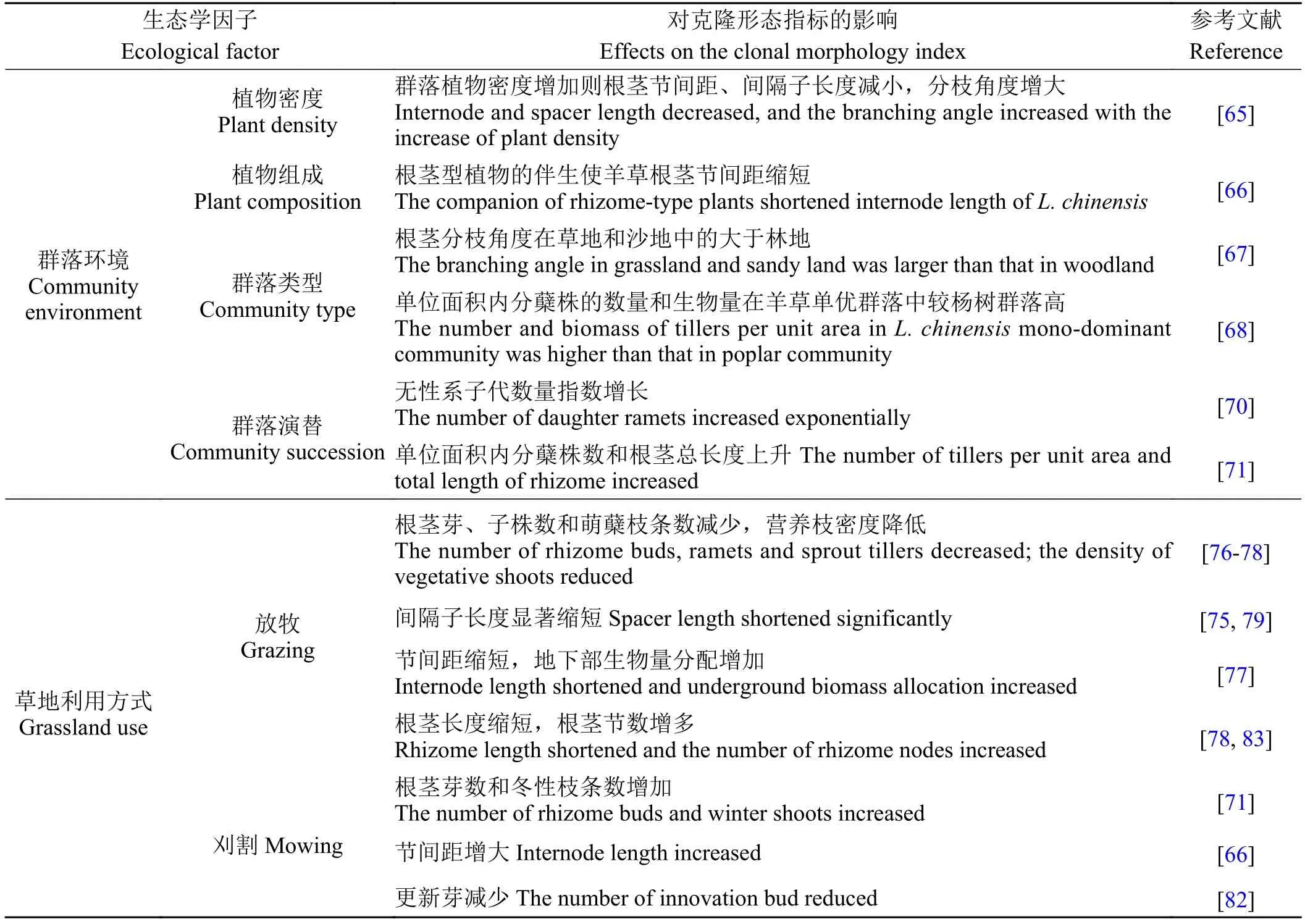
表 4 生态学因子对羊草根茎克隆形态的影响Table 4 Effect of ecological factors on the clonal morphology of Leymus chinensis rhizome
可见群落环境对羊草根茎克隆繁殖产生复杂影响;长期过度放牧不利于羊草克隆繁殖,表现为根茎长度指标降低,芽和分株减少;刈割时间和频次也有影响(表4)。
3 展望
羊草根茎克隆形态可塑性不仅在个体和种群水平发挥重要作用,且对群落和草地生态系统产生深远影响。从羊草根茎克隆形态可塑性研究入手,开展羊草大尺度地理适应性研究具有重要意义。为推动羊草等根茎型草地克隆植物研究的进一步发展,亟待解决以下问题:1)简化研究指标,筛选出能够表征羊草根茎克隆形态的少数关键指标;2)营养缺乏,尤其是N元素和P元素缺乏对羊草根茎克隆形态的影响;3)群落环境和人为扰动对羊草根茎克隆形态的影响;4)对大量的相关研究结果进行Meta分析[84],更准确、客观地评估研究结果;5)羊草根茎克隆形态是否与地上部表型一样具有可遗传性;6)羊草根茎对异质性环境资源是否具有主动适应性?7)羊草等根茎型克隆植物影响群落稳定性和物种多样性的机制如何?如何选择材料和设计试验来进行此方面的研究?8)借助现代科学技术,结合生理和分子方面的研究[85],揭示羊草通过根茎可塑性来适应不同逆境的内在机理。
猜你喜欢
昆明医科大学学报(2022年1期)2022-02-28
昆明医科大学学报(2021年8期)2021-08-13
昆明医科大学学报(2021年6期)2021-07-31
现代畜牧科技(2021年4期)2021-07-21
种子(2020年4期)2020-12-22
今日农业(2020年13期)2020-08-24
中成药(2017年3期)2017-05-17
现代畜牧科技(2016年9期)2016-02-21
草业科学(2015年9期)2015-12-11
西南医科大学学报(2015年1期)2015-08-22
