
香菇单核菌株菌丝生长特性分析*
2019-08-13 09:05程爽爽杜双田
中国食用菌 2019年7期
程爽爽,张 姣,杜双田
(西北农林科技大学生命科学学院,陕西 杨凌 712100)
自古以来,香菇[Lentinus edodes(Berk.)Sing.]就有着“山珍之王”、“植物性食品的顶峰”等美誉,不仅营养丰富,还具有降低胆固醇,预防心血管疾病、糖尿病、佝偻病。
随着人民生活水平的提高及保健意识的增强,香菇的市场需求量逐年增大。截止目前,我国已有“庆元香菇”、“随州香菇”、“西峡香菇”、“平泉香菇”4个地理标志香菇产品,基本形成了以“庆元香菇”产地为核心区域的浙江、以“随州香菇”产地为核心区域的湖北、以“西峡香菇”产地为核心区域的河南、以“平泉香菇”产地为核心区域的河北的4大产区。
香菇菌种在香菇产量及品质构成中占有十分重要的位置,尤其在以自然生态环境和传统农艺方式为基础发展起来的我国香菇产业中,优良菌种在产业发展所占的技术份额起到举足轻重的作用。我国之所以能在几年时间迅速发展为产量居世界第一的食用菌生产大国,除了因地制宜地采用适合中国国情的栽培体系和栽培方式以外,重视引进及选育适合我国栽培特点的优良菌种无疑也是一个重要因素。
我国香菇产量占食用菌总产量的70%。但在香菇的育种方面,与先进国家相比还有一定的差距。我国香菇的育种经历了起步阶段、引种阶段、单孢杂交育种阶段及原生质体技术选育菌种4个阶段[1],主要方法有人工选择育种、杂交育种、诱变育种、原生质体融合育种、基因工程育种、分子标记辅助育种等。其中,杂交选育是目前选育香菇新品种最有效的方法[2]。香菇的杂交育种是利用香菇的担孢子有不同“性”的特性,选择不同优良亲本的单核菌株进行配对杂交以实现基因重组,综合其两亲本的优良性状,克服亲本原有的不良性状,从而育成符合人们需要的、具有杂种优势的新品种。在杂交育种中,选择优良单核菌株是获得优良杂交株的关键,但关于单核菌株生长特性的研究甚少。
本研究通过分析香菇不同品种单核菌株的菌丝生长速率、菌落类型频率分布、菌落长势评分等指标以及低温保藏对单核菌株生长的影响,比较不同香菇品种单核菌株生长性状的差异性,以期为香菇杂交选育的材料选择提供参考。
1 材料与方法
1.1 材料
1.1.1 品种
供试品种见表1。

表1 供试品种编号及其来源Tab.1 Test varieties and their sources
1.1.2 综合PDA培养基
马铃薯200 g、葡萄糖10 g、蔗糖10 g、蛋白胨2 g、酵母浸粉 2 g、KH2PO41.0 g、MgSO4·7H2O 0.5 g、琼脂粉12 g,水1 000 mL。
1.2 方法
1.2.1 单核菌株的获取
参照参考文献 [3],将单孢分离获得的单菌落菌株接至综合PDA平板,培养10 d,经插片镜检,菌丝上无锁状联合即为单核菌株,即为研究材料。
1.2.2 单核菌株菌丝生长速率测定
将上述单核菌株转接至综合PDA平板,在(25±1)℃培养12 d,沿菌落边缘用直径5 mm的无菌打孔器取其菌饼再转接在综合PDA平板中央,记录菌丝萌发时间,采用十字划线法[4]测定菌丝生长速率(S,mm·d-1)公式为:

式中:D为菌落平均直径(mm),T为培养天数(d)。
1.2.3 菌落长势评分
根据单核菌株菌落的洁白程度、气生菌丝的疏密及旺盛程度等将试验菌落分为5组(优、良、中、较差、差),从差到优依次给1分~5分,不萌发菌株的长势评分为0。
1.2.4 菌落类型
在综合PDA平板培养中,单核菌丝较纤细,分枝少,菌丝生长速率缓慢。菌落形态可归纳为4种类型[5]:疏松型(A)、致密型(B,与双核菌丝菌落相似)、贴生不均匀型(C)、贴生均匀型(D),如图1所示。分别统计不同品种的单核菌株不同菌落类型的分布情况。

图1 单核菌株菌落的类型Fig.1 Colony types of monokaryons strains
1.2.5 低温保藏对单核菌株菌丝生长情况的影响
选取4℃低温保藏3个月的单核菌株,在综合PDA培养基上活化2次,依据方法1.2.2转接综合PDA平板,统计菌丝萌发时间,测定菌丝生长速率。
1.3 数据处理
采用Excel 2003和SPSS 20进行分析。
2 结果与分析
2.1 单孢分离及单核菌株鉴定
分离鉴定结果见表2。

表2 不同品种单孢分离及单核菌株数量统计Tab.2 Number of single colonies and number of monokaryons in different strains
由表2可见,研究中挑取的单菌落较多,但得到的单核菌株有限,表明孢子在无菌水中的分散性较差,部分孢子没有完全分开,在今后的研究中可在制备无菌水时添加一定的表面活性剂,使孢子充分分散,以取得更好的分离效果。
2.2 单核菌株菌丝生长速率比较
不同品种单核菌株的菌丝生长速率测定结果见表3和图2。

表3 不同品种单核菌株的菌株生长速率Tab.3 Mycelium growth rates of monokaryonsin in different varieties

图2 不同品种单核菌株菌丝生长速率频率分布Fig.2 Distribution on mycelium growth rates of monokaryons in different varieties
由表3可见,不同品种的香菇,其单核菌株的菌丝生长速率差异显著。其中,L-02的单核菌株菌丝生长速率最小,为0.77 mm·d-1;0912的单核菌株生长速率最大,为2.02 mm·d-1,是L-02的单核菌株的2.62倍;二者的单核菌株菌丝生长速率均与其它品种的差异显著。标准差的大小反映了同一品种不同单核菌株菌丝生长速率差异情况,标准差越大,表明不同单核菌株生长速率差异越大。可以看出,韩香的单核菌株生长速率差异最大,L-02的单核菌株生长速率差异最小。
由图2可知,供试品种单核菌株生长速率分布接近正态分布,但不同品种的分布图形态差异较大,表明不同品种的单核菌株生长速率差异程度不同。图2Ⅰ中,L-02所有单核菌株生长速率均不超过1.50 mm·d-1,主要集中于 0.50 mm·d-1~1.00 mm·d-1,该品种不同单核菌株的生长速率差异性小,且生长速率小,与表3结果相同。韩香、K95-1、K95-2、K95-5、永丰1号的单核菌株生长速率分布较分散,表明该品种单核菌株生长速率差异性较大。图2Ⅳ所示的3个品种,即段木1号、9319和0912,生长速率在0~1.50 mm·d-1之间的单核菌株很少,3个品种的单核菌株生长快的居多,平均生长速率高于其他品种。
2.3 单核菌株菌落长势评分比较
单核菌株菌落长势评分比较见图3。由图3可见,供试品种的单核菌株菌落长势评分频率分布接近正态分布,且集中分布在3分区域,分布频率均在40%以上。表明香菇的单核菌株菌落长势接近均值的居多,长势差的单核菌株数量相对较少。供试品种中大部分品种单核菌株菌落长势相对较好。仅在L-02、康源3号、灵仙1号、段木1号4个品种中,有少部分单核菌株的菌落长势差,但其数量均不超过总样本的6%。灵仙1号和永丰1号单核菌株菌落长势较差的较多,分别为总样本数的16.98%和16.13%。0912单核菌株菌落长势中等的最多,分布频率为77.19%。段木1号和武香1号菌落长势良好的单核菌株较多,分布频率分别为31.53%、31.25%。永丰1号菌落长势优秀的单核菌株最多,分布频率为16.13%。
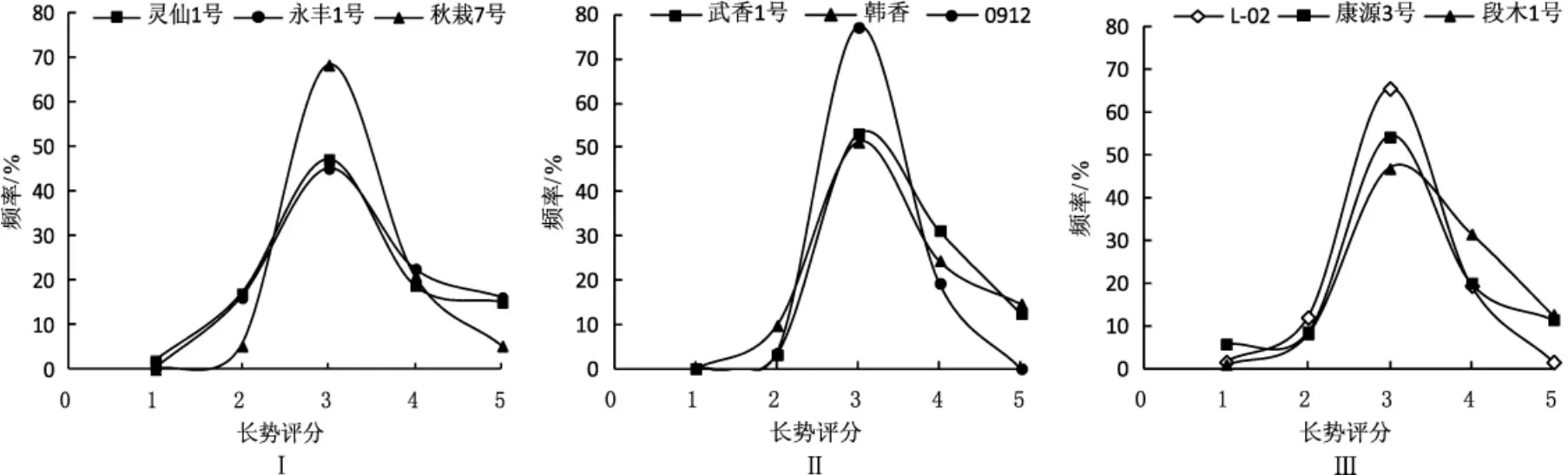
图3 不同品种单核菌株菌落长势评分频率分布Fig.3 Distribution on colony growth scores of monokaryons in different varieties

图4 不同品种单核菌株菌落类型分布Fig.4 Distribution on colony types of monokaryons in different varieties
2.4 不同单核菌株菌落类型分布分析
不同单核菌株菌落类型分布分析见图4。
由图4可知,不同香菇品种单核菌落类型分布差异较大。在4种类型中,疏松型单核菌株分布频率差异最大,其中L-02中疏松型单核菌株占86.76%,段木1号中疏松型仅占9.48%,相差9.15倍;致密型单核菌株分布频率差异次之,其中段木1号中致密型单核菌株占49.14%,L-02中致密型占5.88%;贴生均匀型单核菌株菌落类型差异更小,其中康元3号中贴生均匀型的单核菌株分布频率最大为36.17%,L-02的最小为2.94%;贴生不均匀型单核菌株分布频率差异最小,灵仙1号贴生均匀型的单核菌株分布频率最大为32.08%,L-02的最小为4.41%;同一品种的单核菌株菌落4种类型的分布频率也存在一定差异性,韩香差异最小(20.59%~32.35%),L-02差异性最大(2.94%~86.76%)。
2.5 低温保藏对单核菌株菌丝生长的影响
2.5.1 低温保藏对单核菌株萌发期的影响
低温保藏对单核菌株萌发期的影响见图5。
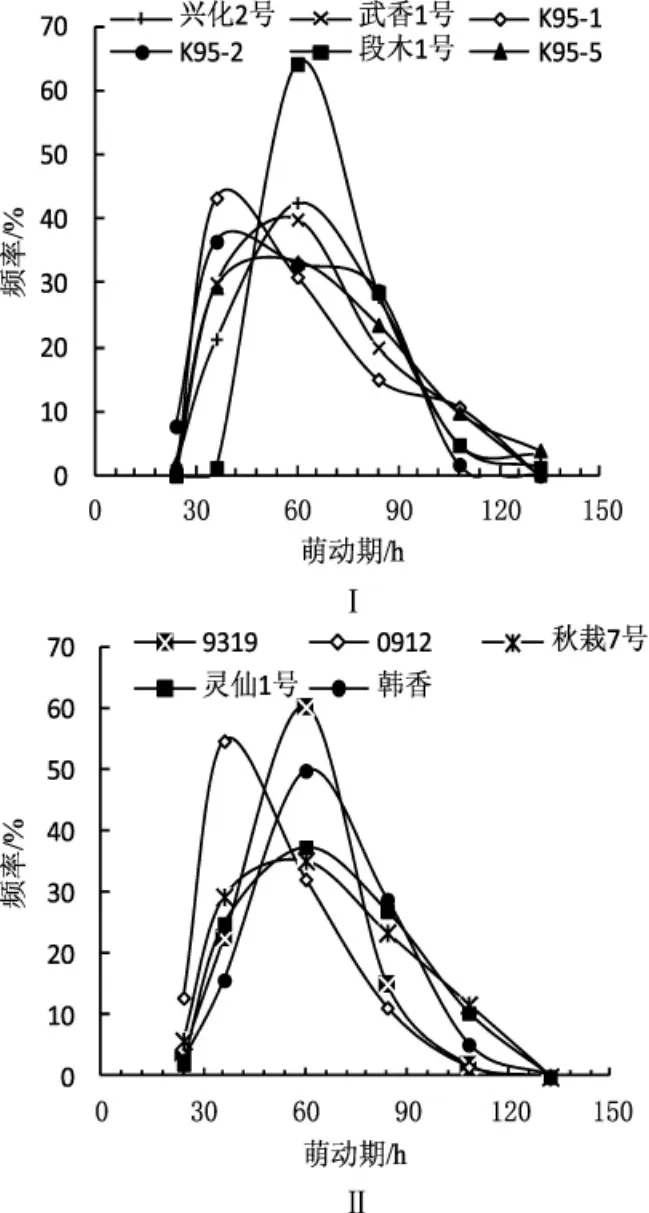
图5 低温保藏3个月单核菌株萌动期分布频率Fig.5 Distribution on germination time of monokaryons
供试单核菌株未低温保藏前其萌动期差异均在24 h内萌发。由图5可知,保藏后的单核菌株萌动期从24 h到132 h不等,低温保藏3个月以后,大部分菌株的萌动期延长。不同品种单核菌株萌动期分布频率差异很大,但都接近正态分布。图5Ⅰ中,段木1号、K95-1、武香1号单核菌株低温保藏后24 h内均不萌发(从图5Ⅰ中看出,3个品种的单核菌株在24 h~36 h范围内均有萌发),其他品种24 h内有少量单核菌株萌发,其中0912最多,为总数的12.90%。0912、K95-1、K95-2的单核菌株在36 h左右萌发较多,段木1号、武香1号、L-02、9319、韩香、灵仙1号、秋栽7号的单核菌株在60 h左右萌发的较多。图5Ⅰ中,段木1号、兴化2号、K95-5少数单核菌株的萌动期为132 h,是低温保藏前萌动期的5.5倍。总体而言,低温保藏3个月使得大部分单核菌株萌动期延长,表明低温保藏可导致单核菌株的活力下降。
2.5.2 低温保藏对单核菌株菌丝生长速率的影响
不同品种单核菌株低温保藏后的菌丝生长速率情况见表4。
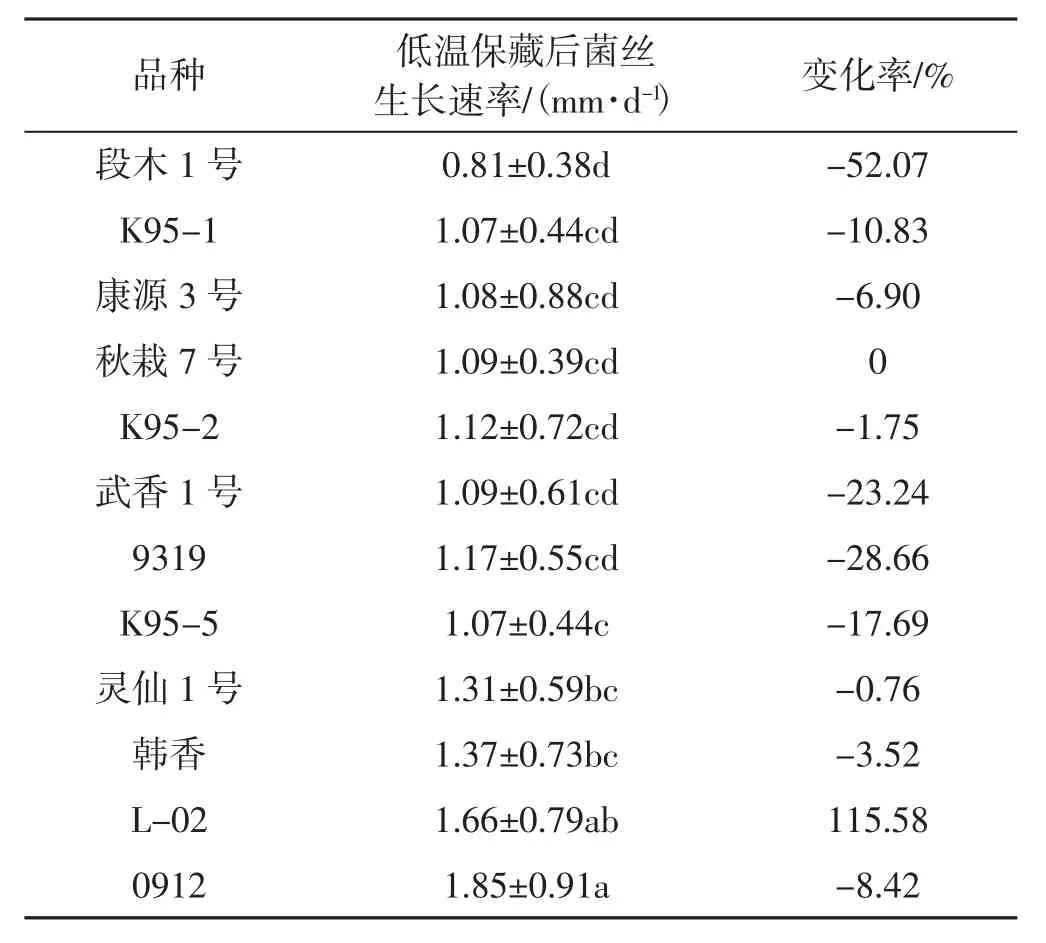
表4 不同品种单核菌株低温保藏后的菌丝生长速率Tab.4 Mycelium growth rates of monokaryons in different varieties after cryopreservation
由表4可见,3个月的低温保藏期对单核菌株的生长速率影响很大。L-02的单核菌株的菌丝生长速率显著提高,是未保藏前的1.15倍。秋栽7号单核菌株的生长速率没有受到影响。其余品种单核菌株的生长速率均有降低。其中,段木1号的下降率为52%,其次为武香1号及9319,再次为K95-5和K95-1等。由此可见,大部分香菇品种的单核菌株不宜长期低温保藏,在单孢分离之后立即进行杂交,以免影响杂交效果。
3 讨论
3.1 不同香菇品种单核菌株生长速率间差异显著
从结果分析看出,供试13个品种单核菌株菌丝生长速率差异显著。0912的单核菌株生长速率最高到达(2.02±0.48) mm·d-1,L-02的单核菌株菌丝生长速率最低为(0.77±0.19) mm·d-1。因为品种不同,其遗传物质必然不同,菌丝生长速率有一定差异。因此在选择单核菌株时不能一味地选择菌丝生长快的菌株,林范学等[6]育种时发现,菌丝生长太快的菌株,基质营养消耗过快,香菇的产量不高,产量主要集中在第一潮、第二潮菇。Gong等[7]对157个香菇单孢单核体的生长速度等性状进行研究,发现菌丝生长速度、生物量、萌发时间呈现连续变异,属多基因控制,生长速度与生物量呈正相关,萌发时间与生长速率、生物量呈负相关。
3.2 同一品种单核菌株菌丝生长速率分布频率不同
同一香菇得到的单核菌株菌丝生长速率间有较大差异,其生长速率的分布频率呈正态分布,表明同一子实体所产生的孢子其遗传因子有一定差异,担孢子间有较大的选择性。从频率分布的标准差来看,其选择差不同。潘迎捷等[8]研究发现,同一来源的香菇单核菌株之间存在酯酶同工酶等酶的多型性,表明同一来源单核菌株遗传物质差异较大。因此采用单孢分离杂交选育是提高菌株生产性能的重要途径。
3.3 单核菌株菌落长势评分符合正态分布
菌落长势是在菌种选择中使用的一个综合性概念,是菌落大小、白度、菌丝旺盛程度等指标的一个综合指标。菌落长势评分符合正态分布表明大部分菌株的生长情况集中在平均数附近,表现差的和优秀的仅占少数,表明以菌落长势为指标选择菌株时,必须有足够的样本数量。
3.4 不同品种单核菌株菌落类型分布频率不同
同一朵香菇的单孢子有4种类型,在分离培养中,单孢子萌发的单核菌株在综合PDA培养基上生长特征不同,分成疏松型、致密型、贴生不均匀型、贴生均匀型4种类型目的是选择不同类型的单核菌株杂交。结果发现,不同的香菇品种中,单核菌株的4种类型所占比例并不一致。在供试品种中,品种9319、L-02和秋栽7号的疏松型单核菌株所占比例较高,分别达到69.49%、86.76%和72.22%,其他类型所占比例相应的就较少。韩香、康源3号、灵仙1号3个品种中4种类型的比例接近。因此,从单核菌株的菌落类型选择杂交组合也有一定的局限性。按照经典遗传学原理,香菇为四极性异核结合,孢子的4种类型不能完全从菌落的生长特性上反映出来。程水明和林范学[9]的研究结果表明,香菇中担孢子交配型因子分离偏离理论预期,偏离程度栽培菌株大于野生菌株。张小雷等[10]认为,单核菌丝挑取的时间导致生长极慢的菌株丢失,进一步造成偏分离现象。所以试验中挑取单菌落的时间可能会导致孢子萌发极慢的菌株丢失,使得菌落类型分布出现偏差,需要调整挑取单菌落的时间进行进一步的试验验证。
3.5 单核菌株不宜低温长时间保藏
研究结果表明,3个月的低温保藏,绝大部分菌株的萌发期延长,菌丝生长速率降低,表明菌株的活力下降。但也有特殊情况,品种L-02的萌发期虽然延长,但生长速率反而增加,这种情况虽不多见,但确实存在,有待于进一步研究。黄年来[5]指出,“分离到单核菌丝后,应立即进行杂交”,“香菇单核菌株随保藏时间的延长也影响杂交成功率”。刘新锐等[11]发现,低温保藏会降低香菇漆酶、羧甲基纤维素酶木糖醇酶和淀粉酶等酶的活力。衣川坚二郎等报道,从香菇得到的单核菌株杂交后都能正常的双核化,将这些单核菌株保存每年转接1次到2次,经几年后,用其中的一些杂交发现22个亲和组合中有3个为正常的双核化,有7个为有限的双核化,有12个为单侧双核化,大部分单核菌株随保藏时间的延长失去了杂交能力。
香菇为四极性异宗结合的担子菌,担子产生4种交配型的孢子,在减数分裂产生孢子的过程中,来自2个核的16条染色体的基因发生重组,导致其携带的遗传物质差异显著。李莉云等[12]发现来自同一菌株的香菇单核菌株不仅有菌丝形态及交配型的差异性,而且存在酯酶同工酶差异。谭琦等[13]利用RAPD技术对香菇孢子单核体的遗传差异变化进行分析,结果表明,同种交配型的孢子单核体遗传相似性分别为66.3%和71.1%,远低于原生质体单核体的。由此可见,香菇在减数分裂过程中染色体交换频率较高,利用杂交选育香菇优良菌株的潜力很大[14]。深入研究单核菌株及其杂交双核菌株的性状,将对香菇的杂交选育工作具有重要意义。
猜你喜欢
今日农业(2022年1期)2022-11-16
今日农业(2022年16期)2022-09-22
新疆农业科学(2022年5期)2022-06-15
陕西农业科学(2022年2期)2022-03-28
食用菌(2021年3期)2021-06-03
浙江农业学报(2021年5期)2021-05-31
陕西农业科学(2021年12期)2021-02-21
中国食用菌(2020年11期)2021-01-18
今日农业(2020年17期)2020-10-27
南京医科大学学报(自然科学版)(2020年2期)2020-03-27
